


| name | Amanita umbrinella |
| name status | nomen acceptum |
| author | E.-J. Gilbert & Cleland |
| english name | "Australian Umber Amanita" |
| images |
 1. Amanita umbrinolutea, eastern Australia. |
| intro |
The following is largely based on the original description (1941). |
| cap |
The cap of Amanita umbrinella is around 75 mm wide, hemispheric at first, convex becoming planar, center becoming depressed, viscid to dry with a lightly striate margin that is sometimes not apparent at first. The cap can range from grayish brown to saturated olive-chestnut, bistre, very dark umber, chocolate, drab hair brown, or olivaceous-black. The volval remnants on the cap are grayish-white warts sparsely distributed or, occasionally, as a single patch (white at first and graying). |
| gills |
Gills are rather crowded, very close to the stem or even attached to it, white or very pale cream colored, often becoming pinkish, and, in dried material, sometimes becoming purplish. The short gills are numerous, squarely truncate, and sometimes seeming to reflect a purplish tint. |
| stem |
The stem is about 75 x15 mm, narrowing upward, with a fibrillose surface. The stem may be more pallid than the cap, but the basal part will often be distinctly gray or gray-brown. The bulb is 25 mm wide and is pronounced or marginate or neither. When the bulb is not clearly defined, the stipe base is sometimes pointed. On the bulb's border, there is a limbate volva which is submembranous to friable and often falling away. The ring is membranous, ample, and persistent. It's white to somewhat sordid and finely striate above. |
| spores |
The spores measure approximately 11.5 - 13 × 9 - 11.5 µm. Presence or absence of clamps on bases of basidia not known. Its author reports that spores were not found in the original material of A. bambra Grgur. Wood (1997) presents a confusing picture on basidial clamps in this species indicating that they can be present and not rare in some cases as well as absent in others. Moreover, he applies the name umbrinella to a taxon with pale pigments. Considering the phenetic similarity of the present species with taxa bearing plentiful clamps, I'm inclined A. umbrinella will be found to bear clamps on its basidia. Very often one can get a small sample of spore size and shape from the illustrations of Gilbert (1941). In the case of A. umbrinella, there is so much variation in the spore shapes drawn for the species that we have chosen only to use the information from the holotype and one paratype (called syntype by Gilbert). Among these only four were presented in lateral view; and (as is our usual microscopic procedure) these were the only spores on which we took data. The resulting measurements yield 11.5 -14 × 8.8 - 9.8 µm and are ellipsoid. |
| discussion |
This species is originally described from the state of South Australia where it was found in sandy soil. Reid (1980) cites reports of its presence in the states of Victoria and West Australia. The authors indicated no nearby tree species. Amanita umbrinella is also reported from New South Wales with Eucalyptus.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita umbrinella | ||||||||
| author | E.-J. Gilbert & Cleland. 1941. Iconogr. Mycol. (Milan) 27: 273, pl. 23 & 24. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Australian Umber Amanita" | ||||||||
| synonyms |
≡Amanitaria umbrinella E.-J. Gilbert & Cleland nom. nud. 1940. Iconogr. Mycol. (Milan) 27: 77, tab. 20 (figs. 3-6), tab. 21, tab. 22 (fig. 1).
=Amanita pantherina sensu Cleland & Cheel. 1914. Agric. Gaz. N. S. Wales 25(10): 887, pl. 1 (figs. 1-2).
=Amanita bambra Grgur. 1997. Larger Fung. S. Austral.: 389, fig. 259(a-d). [Since Grgurinovic does not mention A. umbrinella in her revision of material in AD, this suggests that the holotype of the present species was retained by Gilbert and lost with Gilbert's herbarium. Since the description of A. bambra is apparently based on the same notes on fresh material on which A. umbrinella was based, it seems best to assume that the holotype of the latter is lost. The holotype of A. bambra will need to be revised. If it is consistent with the protolog of A. umbrinella and if, indeed, the holotype of A. umbrinella cannot be found, then the holotype of A. bambra should be designated the lectotype or neotype (as appropriate) of A. umbrinella.]
non Amanita umbrinella sensu Singer. 1954. Sydowia 8: 121. [Singer’s concept included at least A. diemii and . morenoi, based upon RET's examination of collections determined by Singer as A. umbrinella.]
non Amanita umbrinella sensu Wood. 1997. Austral. Syst. Bot. 10: 729, fig. 2(a-e). The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| etymology |
umbrinus "umber" [darker shades of red-brown] + -ella [diminutive ending]; hence, "little umber [thing]" bambra - "mushroom" (from unidentified language of original peoples of Australia) | ||||||||
| MycoBank nos. | 284081, 284095, 443363 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes |
A. bambra—AD. A. umbrinella—designated by Gilbert (1940: pl. 20 (fig. 3) accompanying text) as "Cleland n. 22, 14-iv-1913, water color DIC n. 5, Amanita pantherina." | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material not directly from the protolog of the present taxon or the protolog of A. bambra and not cited as the work of another researcher is based on original research of R. E. Tulloss. | ||||||||
| pileus |
from protolog: ca. 75 mm wide,
grayish deep brown to olive-chestnut, initially
hemispheric, then convex and finally planar and even
slightly depressed over disc, dry or barely viscid,
sometimes shiny; context not described;
margin briefly striate; universal veil
as sparse grayish white warts or
patches. from protolog (A. bambra): up to 102 mm wide, near Drab but a little lighter or darker or Hair Brown or Bister or darker than Saccardo's Umber; context not described; margin not described; universal veil as grayish white patches. | ||||||||
| lamellae |
from protolog: barely adnate, moderately crowded, white to pale cream, often taking on pinkish tint in age, becoming more or less distinctly purple in drying; lamellulae squarely truncate. from protolog (A. bambra): nearly free to adnexed, moderately close, white to cream colored, up to 6 mm broad; lamellulae not described. | ||||||||
| stipe |
from protolog: ca. 75 × 15 mm, white, tinged with color of pileus below, robust, cylindric or slightly narrowing upward, fibrillose; bulb up to 25 mm wide,occasionally pointed [below]; context solid or stuffed then hollow; partial veil membranous, ample, white or rather sordid, lightly striate, persistent; universal veil as free margin or limbate volva, friable, fugacious. from protolog (A. bambra): up to 89 × 32 mm, whitish, sometimes grayish above(?), "swollen above," slightly striate above; bulb ?; context ?; partial veil "median to distant, slight to ample", striate above, "soon disappearing"; universal veil "at first with free edge," friable, evanescent. | ||||||||
| odor/taste | not described. | ||||||||
| macrochemical tests |
none described. | ||||||||
| pileipellis | not described. | ||||||||
| pileus context | not described. | ||||||||
| lamella trama | not described. | ||||||||
| subhymenium | not described. | ||||||||
| basidia | from protolog (A. bambra): [15/1/1] 46 - 62 × 9.6 - 14.4 μm, avg. length = 51 μm, avg. width = 12.3 μm, 4-sterigmate, with sterigmata up to 5.6 μm long; clamps absent. [Note: Because of the similarity of the present species with A. murinoflammeum, it is probable that clamps will be found at the bases of basidia in the present species.—ed.] | ||||||||
| universal veil | from protolog (A. bambra): On pileus: filamentous hyphae 2.0 - 5.6 μm (avg. = 3.9 μm), occasionally branched; inflated cells ovoid or clavate or obpyriform. | ||||||||
| stipe context | not described. | ||||||||
| partial veil | not described. | ||||||||
| lamella edge tissue | from protolog (A. bambra): inflated cells clavate or obpyriform or subglobose. [Note: This tissue, described by Grgurinovic as "veil fragments," appears to correspond, at least in part, to lamella edge tissue.—ed.] | ||||||||
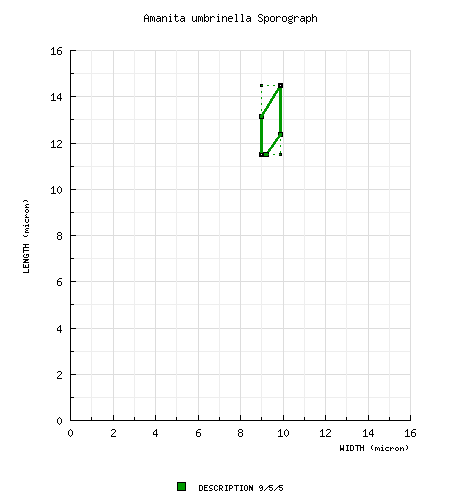
| basidiospores |
from protolog (spores of type): [2/1/1] 10.2 × 8.0 μm, (Q = 1.28), hyaline, smooth, inamyloid, broadly ellipsoid; apiculus sublateral (per figure); contents not described; white in deposit. [Note: The spore measurements are derived from measuring scale drawings of spores of the holotype (Gilbert 1940: tab. xx(fig. 3)) using only spores that are presented in lateral view.—ed.] from protolog (spores from all original material): [9/5/5] 11.5 - 14.5 × 9.0 - 9.9 μm, (L' = 12.6 μm; W' = 9.3 μm; Q = 1.25 - 1.46; Q' = 1.35), hyaline, smooth, inamyloid, broadly ellipsoid to ellipsoid. [Note: The spore measurements are derived from measuring scale drawings of spores of original material (Gilbert 1940: tab. xx(figs. 3-5) and xxi(figs. 1 & 3)) using only spores that are presented in lateral view. The spores of tab. xxi(fig. 5) are excluded because they are so much smaller than those in the remaining figures (e.g., 9.0 × 6.8 μm).—ed.] from protolog (A. bambra): [56/-/11] 9.2 - 14.2 (-16.8) × 7.2 - 11.4 (-12.8) μm, (L = 11.7 μm; W = 9.5 μm; Q' = 1.20), "subglobose" to broadly ellipsoid, amyloid, adaxially flattened (per figure); apiculus sublateral, cylindric (per figure); contents not described; color in deposit not recorded. [Note: We have not estimated a range for Q based on the (Grgurinovic 1997) spore data for this species in order to prevent automatic generation of a sporograph that we believe would be misleading. We evaluated the Amanita spore length and width ranges from (Grgurinovic 1997) in comparison to comparable data published by other authors and often based on revision of the same specimens. This experiment involved a total of 19 descriptions of a total of 13 species from the work of 3 authors. In a range of the form "x - y" of spore length (width) from (Grgurinovic 1997) compared to a range of the form (a-) b - c (-d) of spore length (width) in the other works, the value of "y" was compared to the value of "c" as a ratio. In the case of spore length ranges, on average (per author), the ratio y/c ranged from 1.06 - 1.10 (possibly due to the non-segregation of a "d" value in the ranges of concern). In the case of spore width ranges, on average (per author), the ratio ranged from 1.14 - 1.23 (indicating the probability of compounding causes at play—possibly, the absence of the "d" value in the ranges of concern and failure to restrict spore measurement to spores strictly presenting in lateral view). When sporographs were attempted from the Grgurinovic data, the results were not useful.—ed.] | ||||||||
| ecology | from protolog: Terrestrial in sandy soil. | ||||||||
| material examined |
from protolog: AUSTRALIA: NEW SOUTH WALES—Unkn. LGA—Milson Isl., Hawkesbury R., 14.iv.1913 unkn. coll. [J. B. Cleland? 22] (holotype, in herb. Gilbert, as "A. pantherina" & exsiccatum lost & watercolor reproduced in (Gilbert 1941: tab. 23)). [Note: Gilbert lists 11 additional collections determined by him as A. umbrinella, but with very limited information. It is possible they were all lost with Gilbert's herbarium. They are presented in the order in which their spores are figured in tabs. xx - xxii. Reid (1980) states that all the collections came from New South Wales or South Australia. This is the best we can do at present in terms of identifying them: ] from protolog (A. bambra): AUSTRALIA: SOUTH AUSTRALIA—LGA Unkn. - Adelaide, Beaumont Common, 19.vii.1925 unkn. coll. s.n. (paratype, AD 3036, also paratype of A. umbrinella); Belair Nat. Pk., 6.viii.1921 unkn. coll. s.n. (paratype, AD 3047); Dashwoods Gully, 10.x.1919 unkn. coll. s.n. (paratype, AD 3059); Kinchina, 1.viii.1925 unkn. coll. s.n. (holotype, AD 3050); Mt. Compass, 26.x.1920 unkn. coll. s.n. (paratype, AD 3060); Mt. Lofty, 7.iv.1917 unkn. coll. s.n. (paratype, AD 3063); Morialta, 3.vi.1933 unkn. coll. s.n. (paratype, AD 3042); Myponga, 26.v.1926 unkn. coll. s.n. (paratype, AD 3038, possibly also paratype of A. umbrinella), 10.x.1929 unkn. coll. s.n. (paratype, AD 3035); Stirling West, 23.vii.1927 unkn. coll. s.n. (paratype, AD 3034). | ||||||||
| discussion |
This taxon has been described twice. Both
times the descriptions were based on material
collected by J. B. Cleland and associated field
notes, sketches, photographs, and at least one or
two watercolors. One collection appears to
have served as paratype for both names; and another
collection may have. Gilbert specifically
states that his description is based on his choice
of holotype and its accompanying water color.
He then provides the full set of reported, variant
colors of the pileus in his discussion.
Considering the degree of variability in pileus
coloration and the very
variable shape of spores drawn from the collections
that were in the possession of Gilbert, it would
have been valuable if Grgurinovic had similarly
restricted the description of A. bambra to
her choice of its holotype. However, this was
not the case. Hopefully, the holotype of A.
bambra is a collection for which there are good
field notes and, possibly, a painting, sketch, or
photograph. From material sent by Cleland, Gilbert extracted a set of collections from the mixed group that Cleland had assigned to his mistaken concept of A. grisea. Cleland himself had noted how difficult it was to deal with this group and had struggled to subdivide it in some way. Gilbert extracted the amyloid-spored A. grisella (sect. Validae) from the same cluster of collections. He remarks that the macroscopic descriptions of fresh material of the two taxa made for a confusing situation. He states that he was able to describe the type collection of A. umbrinella thanks to the existence of a watercolor illustration of it [reproduced by Gilbert (1941: tab. 23)]. Unfortunately, we don't know what characters of the type and its watercolor Gilbert used in order to determine the conspecificity of the paratypes. However, the result, as Gilbert notes, is a "rather polymorphic species" with, for example, the tint of the cap varying from Bister to Chocolate to Drab to Hair Brown and Dark Olive. Likewise, Gilbert notes the basidiomes may be either robust or slender; and the bulb of the stipe may be pronounced or not and may be marginate or not. The picture is further complicated by proposal of the name A. bambra (Grgurinovic 1997) as noted, above. The set of field notes from which Grgurinovic presumably operated in writing the macroscopic description of A. bambra can be seen to have a great deal of overlap with regard to the data provided by Gilbert (1941). We don't know, at present, exactly what part of the protolog of A. bambra comes from descriptive materials and/or illustrations associated with the holotype. If Gilbert and Grgurinovic are correct in the assumption that they were each describing a single (although apparently polymorphic) species, then there is no problem. If it were to be demonstrated that A. umbrinella and A. bambra are species clusters, then it may be the case, by chance, that the names are taxonomic synonyms or that they apply to two different species within the hypothetical cluster. To be prepared for any eventuality, it will be necessary to know the content of extant, supportive materials for the holotype of A. bambra. RET believes that there should be real concern over whether there are multiple taxa involved in Cleland's collections assigned to A. umbrinella and A. bambra by their authors. The basidiome on the right in Gilbert's tab. 23 has pallid orangish cream lamellae, a different partial veil from the dark capped basidiome on the left, a limbate or saccate volva, and an apparently totally elongating stipe. This image could very well represent A. roseolamellata of sect. Caesareae. Should this plate be a candidate for lectotype of A. umbrinella (see below), then the designation should explicitly exclude the species on the right because of the above observation. It is worth note that the one laterally aligned spore of Gilbert's tab. xxi(fig. 5) [excluded from use in the basidiospore data field (above)] has width and Q values that fall within the current known ranges for A. roseolamellata. The following figure provides comparison of the sporographs from this page with the sporograph of A. murinoflammeum a very similar species. ...more... t.b.d. Lectotypification proposal The herbarium of Gilbert is presumed lost and, with it, the holotype of A. umbrinella. At the very least, it would be worthwhile to lay out the argument for how one might proceed in lectotypification of A. umbrinella. A stake in the ground would be useful at this point. There is no known isotype for A. umbrinella. The set of collections selected for original material of A. bambra does not include any material that is a possible isotype for A. umbrinella. Because Gilbert must be credited with segregation of A. umbrinella from Cleland's material, only material seen by Gilbert can be considered original material. Hence, the only known exsiccata that could serve as lectotype are the collections in AD that are possible duplicates of paratypes listed in Gilbert's captions on spore drawings—AD 3036 has the same collection locality and date as the specimen Gilbert cites as "n. 25", and AD 3038 has the same date as one of the two specimens Gilbert cites as "n. 24." If the two collections in question were indeed divided at some time, they were divided by Cleland, who had not formed a concept allowing him to reliably separate the taxa within his concept of "A. grisea." Hence, with regard to possible duplicates in AD, we can have no certainty that the material in those collections is conspecific with the material Cleland sent to Gilbert. I would argue that the duplicates in AD must be excluded from the set of original material. If the exsiccata are judged to be questionable with regard to classification as original material, then there remain the published illustrations in (Gilbert 1940 & 1941)—a photograph, a watercolor, and 12 sets of spore drawings. The photograph provides no color information, which seems to me a critical flaw in this case; moreover, there is notable variation in the form of the stipe's bulb in the three specimens depicted; since there is reason to be suspicious that the "polymorphism" of Gilbert's concept hid within it two or more separate species, the photograph seems an inappropriate choice for lectotype. The watercolor and one set of spore drawings are linked to the holotype collection by Gilbert. The spore drawing set contains three spores of which one (or possibly two) are in lateral view; however, standing alone, these drawings are like the spores of many other species of Amanita and, additionally, represent a less than minimal sampling. We are left with the watercolor as a candidate for the lectotype—that is to say, the basidiome depicted on the left of Gilbert's tab. 23 (see above). Gilbert describes the difficulty of dealing with possible mixed collections and explicitly attributes his basic conception of the species to the lost holotype and this painting: "A cause de l'apparence, ces deux amanites se trouvaient confondues, et dans les notes de récolte, il est dificile de savoir si les caractères descriptifs, notés sur le frais, s'appliquent à l'un ou à l'autre espèce. J'ai cependant pu établir la description du A. umbrinella d'après la récolte type, d'après laquelle a été peinte l'aquarelle." (Gilbert 1941: 273-274) {These two amanitas [A. umbrinella and A. grisella] were found to be confused based on their [similar] appearance; and, in the collectors' notes on fresh material, it is difficult to know to which of the two species the descriptive terms are applied. However, I was able to write the description of A. umbrinella based on the type collection from which the watercolor was painted.}In addition to the above argument, which RET judges to be sufficient, it also seems wise to select a lectotype illustrating a very dark brownish gray or brownish black cap because, it appears that contemporary usage of the name in Australia includes application to such material as is depicted in the illustration on this page. Presuming the watercolor to be lost, it then seems most reasonable to designate the image of the leftmost basidium of tab. 23 of (Gilbert, 1941) as the lectotype of A. umbrinella. This designation is made provisionally while RET awaits comments on the proposal. | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.