


| name | Amanita diemii |
| name status | nomen acceptum |
| author | Singer |
| english name | "Diem's Amanita" |
| intro |
|
| cap |
Amanita diemii has a cap 33 - 78 mm wide that is initially bright golden yellow to orange-ocher with the center yellow. Over time, the colors progressively darken through various tones to coffee or burnt umber with the margin always paler. It is convex at first, and often becomes depressed in the center or concave in age. It lacks an umbo, is always viscid, and often appears to bear fine scales or "areolate squamules" (sometimes teak colored) over the center (and sometimes with smaller squamules toward margin)—although the cap dries smooth. The cap''s flesh is largely white, yellow under pileipellis (at least at first), and unchanging when cut or bruised. The margin of the cap is short sulcate at first, then sulcate (to about one-quarter of the cap radius) or long pectinate, never uneven, often decorated with tooth-shaped fragments of volva. The volval remnants at first are found as broad rather flat patches (sometimes proportionately rather thick) that are white (occasionally drying pale grayish) and fibrillose-felted. Sometimes these patches break up into concentric appressed scales that can be removed. The skin of the cap is separable; but, in older specimens, it often ruptures into small scales in the region between the central area and the marginal striations. The latter is an unusual character in sect. Amanita. |
| gills |
The gills of this species are free, crowded to close, white, rather narrow, with edges that are often serrulate to crenulate or floccose-fimbriate; the short gills are truncate, infrequent(?), and unevenly distributed. |
| stem |
The stipe of A. diemii is 45 - 98 × 8 - 18 mm, white at first, eventually light cinnamon, at first narrowing upward, then becoming cylindric, densely flocculose-farinose to pruinose, and never glabrescent in age or even after heavy rain. The stipe's bulb is usually distinct, 15 - 21 mm wide (14 - 21 × 10 - 17 mm and subglobose to subnapiform to napiform in dried specimens). The flesh of the stem is white, unchanging when cut or bruised, stuffed at first and later hollow. The annulus is often lacking and, when found, is very fragmentary and ephemeral, white, thin, and membranous. The volva is white, felted-membranous, rather thin, and appears as a circumcissile free limb [seeming ocreate--suggesting volva and bulb of A. pantherina (DC.:Fr.) Krombh. if free limb collapsed on stipe base] or as dentate sublimbate margin on the top of the bulb [suggesting the volva of A. gemmata (Fr.) Bertillon in Dechambre]]. |
| odor/taste |
Amanita diemii has no odor and a mild taste. |
| spores |
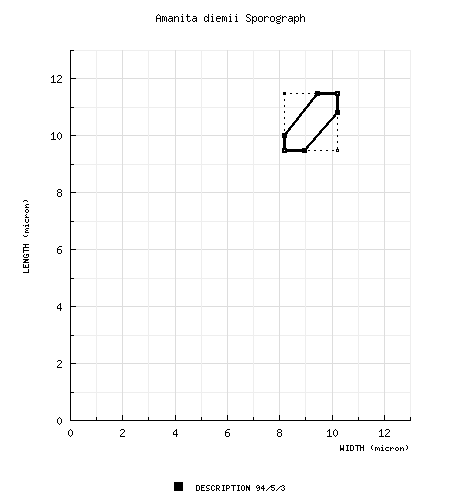
The spores of this species measure (9.0-) 9.5- 11.5 (-12.8) × (7.5-) 8.2 - 10.2 (-11.5) µm and are subglobose to broadly ellipsoid (or infrequently globose) and inamyloid. Clamps are occasionally found at the bases of basidia. |
| discussion |
Amanita diemii is found under Nothofagus in Argentina and Chile. If there is a very similar taxon (none are known, although perhaps the "pantherina-gemmata-group" includes a candidate), such an entity probably also occurs only in the Southern Hemisphere. Older specimens of this species have been misdetermined as A. umbrinella E.-J. Gilbert & Cleland, which is an Australian taxon. The species may be toxic and likely to produce symptoms similar to those of A. muscaria (L. : Fr.) Lam. and A. pantherina (DC. : Fr.) Krombh.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita diemii | ||||||||
| author | Singer. 1954. Sydowia 8: 120. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Diem's Amanita" | ||||||||
| MycoBank nos. | 292447 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | LIL; isotype, BAFC | ||||||||
| type studies | Tulloss, included here | ||||||||
| revisions | Tulloss, here | ||||||||
| intro | [NOTA: RET is not satisfied with his interpretations of oberservation that are made below on this page. This page requires revision.] | ||||||||
| pileus | 33–78 mm wide, initially bright golden yellow to orange-ocher with disc (pl. 11 H 6, H 8; pl. 12 G 8) jonquil (MP) to sunlight (MP), progressively darkening, becoming mummy (MP) to mocha bisque (MP) to santos (MP) to conga (MP) to coffee (MP) to burnt umber (MP), with margin always paler [at first aztec (MP), then pl. 11 G 4 to pl. 10 D 4 to fallow (MP)], convex at first, eventually often with depressed disc or concave, lacking an umbo, always viscid, often appearing verruculose or exhibiting fine scabrosity or "areolate squamules" (sometimes teakwood (MP)) over disc (and sometimes with smaller squamules toward margin), but drying smooth; context white, yellow under pileipellis (at least at first), unchanging when cut or bruised; margin short sulcate at first, then sulcate (0.25R) or long pectinate, never uneven, often decorated with tooth-shaped fragments of universal[?] veil; universal veil at first as broad patches, subapplanate, sometimes proportionately rather thick, white (occasionally drying pale grayish), fibrillose-felted, sometimes eventually breaking into concentric appressed scales, detersile; pileipellis separable, in older specimens sometimes rupturing into small scales in region between disc and marginal striations. | ||||||||
| lamellae | free, crowded to close, white, rather narrow, with edge often serrulate to crenulate or floccose-fimbriate; lamellulae truncate, infrequent(?), unevenly distributed. | ||||||||
| stipe | 45–98 × 8–18 mm, white at first, eventually light cinnamon, at first narrowing upward, then cylindric, densely flocculose-farinose to pruinose, never glabrescent in age or even after heavy rain; bulb usually distinct, 15–21 mm wide (14–21 × 10–17 mm and subglobose to subnapiform to napiform in exsiccata); context white, unchanging when cut or bruised, stuffed then hollow; partial veil lacking or very incomplete (fragmentary) and ephemeral, white, thin, membranous; universal veil white, felted-membranous, rather thin, as circumcissile free limb [appearing ocreate—suggesting volva and bulb of A. pantherina (DC. : Fr.) Krombh. when free limb collapsed on stipe base] or as dentate sublimbate margin on top of bulb. | ||||||||
| odor/taste | Odor lacking. Taste mild. | ||||||||
| macrochemical tests |
Macrochemical tests: HCl - negative. KOH and NH4OH - on pileus cuticle, more vividly tan-ochraceous than surrounding surface. | ||||||||
| pileipellis | 310–505 µm thick in material with sporulation just beginning (holotype), 115–235 µm thick in material with fully expanded pileus (Singer M10539); sponge-like suprapellis (possibly lower portion of universal veil?) of tangled hyphae separated by relatively large gaps 135–185 µm thick in young material, 15–50± µm thick in mature material, probably having been continuous with volva originally, with many broken ungelatinized or partially gelatinized hyphal fragments projecting upward (in cross-section), colorless to yellow near surface (to depth of up to 35 µm), otherwise yellow-orange; compressed subpellis of densely packed, periclinally oriented hyphae 175–340 µm thick in young material, 110–180± µm in mature material, yellow-orange; filamentous, undifferentiated hyphae 2.1–8.8 µm wide, branching, partially gelatinized throughout, commonly with subrefractive walls in suprapellis, occasionally with such walls in subpellis; vascular hyphae not observed. | ||||||||
| pileus context | filamentous, undifferentiated hyphae 2.8–16.0 µm, branching, plentiful to locally dominating, loosely interwoven, singly and in fascicles, occasionally with yellowish subrefractive walls; acrophysalides elongate-ellipsoid to broadly clavate to clavate to narrowly clavate, plentiful to locally dominant, thin-walled, up to 117 × 40 µm; vascular hyphae absent (holotype) or locally common (Singer M10539), 3.5–19.0 µm wide, infrequently branching, sinuous, sometimes coiling tightly. | ||||||||
| lamella trama | bilateral, almost completely collapsed in holotype; wcs = 20–35 µm (poor to moderate rehydration); subhymenial base (as rehydrated) not clearly differentiated from subhymenium, with inflated cells of subhymenial tree subglobose to elongate ovoid to clavate, up to 30 × 20 µm, with numerous filamentous, undifferentiated hyphae giving rise to inflated cells at diverse distances from central stratum, with angle of divergence up to 45°; filamentous, undifferentiated hyphae 2.5–8.2 µm wide, with branching not observable; terminal inflated cells not observed; vascular hyphae not observed; clamps present. | ||||||||
| subhymenium | completely collapsed in holotype; wst-near = 20–25 µm (moderate rehydration); wst-far = 40–50 µm (moderate rehydration); branching structure, with one to three cells between bases of longest basidia/-oles and central stratum (as rehydrated), with three to five cells between bases of shortest basidia/-oles and central stratum (as rehydrated), with basidia arising from clvate to ovoid to subglobose inflated cells often arising directly from filamentous, undifferentiated hyphae. | ||||||||
| basidia | 31–61 × 9.5–14.0 µm, 4- and infrequently 2-sterigmate; clamps infrequent. | ||||||||
| universal veil | On pileus: badly collapsed, partially gelatinized, and infested with mold (with hyphae septate and having yellowish subrefractive walls) in holotype; filamentous, undifferentiated hyphae 1.4–8.8 µm wide, branching, dominating, occasionally anastomosing, occasionally constricted at septa, in fascicles (quite often) or singly, interwoven in open lattice, often loosely coiling; inflated cells scattered to locally common, subfusiform to elongate to narrowly clavate to bacilliform, up to 85 × 15.2 µm, thin-walled, terminal (singly); vascular hyphae rare (?10± µm wide) or absent; clamps locally common. On stipe base: filamentous, undifferentiated hyphae 2.5–10.5 µm wide, branching, dominating, in fascicles and singly, interwoven in rather dense mat, frequently partially gelatinized (holotype), occasionally constricted at septa; inflated cells clavate to narrowly clavate to subfusiform, thin-walled, terminal (singly), scattered, unevenly distributed, up to 57 (–90?)× 18.0 (–20?) µm; vascular hyphae not observed; clamps locally common. | ||||||||
| stipe context | longitudinally acrophysalidic; filamentous, undifferentiated hyphae 2.2– 11.2 µm wide, branching, plentiful; acrophysalides dominating, clavate to allantoid to subcylindric to broadly clavate, up to 295 × 59 µm, with walls thin or up to 0.8 µm thick (especially notable at some apices), occasionally broadly rounded at base so that apex and base difficult to differentiate, occasionally apparently in chains (some appearing dissociated in Singer M10539); vascular hyphae not observed; clamps present. | ||||||||
| partial veil | (only very fragments remaining in holotype) filamentous, undifferentiated hyphae 1.5–10.5 µm wide, branching, mostly in fascicles, with some arrange subradially, but others curving to form open lattice and closely enwrapping inflated cells, occasionally with yellowish subrefractive walls; inflated cells difficult to discern in bundles of hyphae, common?, up to 66 × 35± µm, thin-walled, terminal (singly); vascular hyphae not observed; clamps present. | ||||||||
| lamella edge tissue | not examined. | ||||||||
| basidiospores | [94/5/3] (9.0–) 9.5–11.5 (–12.8) × (7.5–) 8.2–10.2 (–11.5) µm, (L = 10.0–10.7 µm; L’ = 10.4 µm; W = 9.0–9.8 µm; W’ = 9.3 µm; Q = (1.02–) 1.06–1.22 (–1.28); Q = 1.10–1.15; Q’ = 1.12), hyaline, colorless, thin-walled, smooth, inamyloid, subglobose to broadly ellipsoid, infrequently globose, adaxially flattened, often slightly swollen at one end; apiculus sublateral, cylindric, prominent, occasionally proportionately rather broad; contents granular to mono- to multiguttulate with or without additional small granules; pure white ["A (Crawshay)"] in deposit. | ||||||||
| ecology | Subgregarious. Only reported site elev. is 550 m. Argentina: Under Nothofagus dombeyi. Chile: Garrido-G. and Bresinsky (1985) categorize A. diemii as exclusively mycorrhizal with Nothofagus. Garrido-G. (1988) lists the following species as mycorrhizal associates of A. diemii: N. alpina, N. antarctica, N. dombeyi, N. glauca, and N. obliqua. He further lists the following plants as also present at one or another collecting site: Berberis sp., Chusquea quila, Citronella mucronata, Drymis winteri, Eucryphia glutinosa, Laurelia sempervirens, Lomatia ferruginea, and Lophosoria quadripinnata. Singer and Moser (1965: 133) report A. diemii in mixed forest with N. dombeyi and N. alpina. Valenzuela et al. (1999) report that the present species is common in all native Nothofagus forests investigated—with N. antarctica, N. alpina, and N. obliqua as primary symbionts in the Cordillera de la Costa; with N. dombeyi in the Cordillera de los Andes; and with N. dombeyi, N. obliqua, and other unnamed Nothofagus spp. in the Depresión Intermedia. | ||||||||
| material examined | ARGENTINA: NEUQUÉN—Parq. Nac. Nahuel Huapí, Brazo Blest, 29.ii.1988 R. Singer M10539 (F 1075918 as "A. umbrinella"); Parq. Nac. Nahuel Huapí, Quetrihué, 13.v.1952 R. Singer M562 (holotype, LIL n.v.; isotype, BAFC 30.600), 14.v.1952 R. Singer M583 (paratype, LIL? n.v.). CHILE: LOS LAGOS—Valdivia, Cordillera Pelada, "Chivería," 29.iii.1963 R. Singer M3226 (BAFC 30.601; MICH n.v. as "f. macrospora [herb. name]"). | ||||||||
| discussion |
This discussion and the work above are not complete. [NOTA: This "curious suprapellis" is probably better understood as the lower part of the universal veil that remains attached to the pileus.]The curious suprapellis of A. diemii seems to be reduced by age and expansion of the basidiome. The specimens of the holotype are immature or just beginning sporulation. However, the specimen of Singer M10539 is mature—the pileus is fully expanded. In this specimen the thickness of the pileipellis is 115–235 µm thick overall; and the suprapellis is radically reduced in thickness (15–50± µm thick). The scabrous or scurfy nature reported on the disc of the pileus in Singer’s notes on several collections of this species seems to be due to the nature of the suprapellis—especially, the plenitude of broken upstanding hyphae. In his notes, Singer emphasized that the spores of A. diemii were less that 11 µm long; undoubtedly this view was based on the type collection, in which all the basidiomes were immature or just beginning sporulation when dried. Because of this view, he took the Chilean material (Singer M3226) to represent a distinct form. My examination does not support establishment of an additional taxon. [Might be worthwhile to get info. on distribution of N. alpina and N. dombeyii.] On the pileus of the holotype, there is a considerable number of hyphae of a mold infesting the remnants of universal veil. Because the walls of these hyphae are yellowish and subrefractive, it was difficult to be sure whether vascular hyphae were present or not. If they were present, they were very uncommon. Corner and Bas (1962) describe A. elata (Mass.) Corner & Bas as being very similar to the present taxon. .... According to the protolog, when the volval material is lost from the pileus, this species may have the appearance of a species of Russula. | ||||||||
| citations | —R. E. Tulloss and E. Horak | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.