


| name | Amanita maryaliceae |
| name status | nomen provisorum |
| author | Tulloss |
| english name | "Mary Alice's Amanita" |
| images |
  1. Amanita maryaliceae, Mohawk Tr. St. For., W. of Charlemont, ca. border of Berkshire & Franklin Cos., Massachusetts, U.S.A. |
| cap | The cap is 50 - 90 mm wide, white and somewhat translucent, hemispheric to bell-shaped to rather flattened or (occasionally) slightly concave. The cap color does not change when it is cut or bruised. The surface is tacky at first, shiny, with a suggestion of embedded concolorous radial fibrils. The cap flesh is white, with a watery line just above the gills, and does not change when cut or bruised. The cap's nonappendiculate margin is usually not striate, although sometimes it may become faintly striate for about one-tenth of the cap's radius or longer in age. Volval remnants are absent from the cap or may be present as irregular warts or a submembranous patch of loosely connected warts; the remnants are white to off-white, easily crumbling, easily removed, and tend to become grayish with age. |
| gills | The gills of this species are free to narrowly adnate with a decurrent tooth and (often) a decurrent line on the top of the stem. They are white in mass (sometimes faintly pink at first), white in side view, and do not change when cut or bruised. Occasionally to rather commonly adjacent gills may be connected by at some point. Also, the gills may fork, but with open end of the forked gill pointing toward the stem rather than toward the cap's margin. The short gillls are truncate to rounded truncate to subtruncate to subattenuate to attenuate; they are sometimes adjacent to the stipe rather than to the pileus margin; and they are unevenly distributed, of diverse lengths, and plentiful. |
| stem | The stem is 57 - 135 × 5 - 12 mm, white, and does not change when cut or bruised; it is decorated with fine fibers or flocculent material. The stem's bulb measures 22 - 42 × 14.5 - 21 mm; it is somewhat turnip-like to somewhat like a short carrot, with white mycelium threads attached to it. The stem's flesh is solid to firmly stuffed, white, unchanging when cut or bruised or rarely turning pale pinkish brown in the lower part of the stipe. The stem's ring is at or near the top of the stem. It is ample, skirt-like, striate above, and often has a line of beige (eventually graying) volval material on the ring's edge or on its underside near edge. The ring eventually collapses on the stem. The volval may not be evident, or it may be present as an easily detached patch or patches that can accidentally be left in the soil during the collecting process. At other times, the volva may be present in broken collars or irregular rows. In mature material, the volva is off-white to very pale grayish, and often darkens with age. |
| odor/taste | The odor of A. maryaliceae is mild or pleasant when the flesh is cut, close to "floral," or sometimes strongly "floral." The taste has not been recorded. |
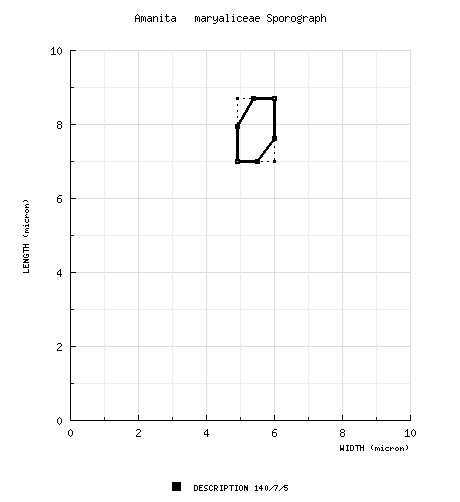
| spores | The spores of this species measure (6.3-) 7.0 - 9.0 (-9.5) × (4.5-) 4.9 - 6.3 (-7.0) µm, are broadly ellipsoid to ellipsoid (or, occasionally, elongate) and amyloid. The basidia lack clamps at their bases. |
| discussion | This species is only know from a very limited range at present. Molecular studies show that the name has been incorrectly applied to Amanita media/A. radiata in the Gulf Coast area and to A. ostendemihi over the range of that species.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita maryaliceae | ||||||||
| author | Tulloss nom. prov. | ||||||||
| name status | nomen provisorum | ||||||||
| english name | "Mary Alice's Amanita" | ||||||||
| etymology |
genitive of a Latinized name; hence, "of Mary Alice" or "Mary Alice's." In honor of RET's partner and wife, Mary [Alice] King Tulloss, who has assisted in preparing his work for publication until she can say "undifferentiated filamentous hyphae" and "longitudinally acrophysalidic" in her sleep. She has collected with him, annotated collections with him, helped to edit the original Amanita Studies website, reviewed DNA annotations and metadata in Sequin files to be submitted to GenBank, carried out filing and other secretarial tasks for him, assisted at his lectures, and (as a retired software developer and a teacher of high school Mathematics) played a significant role in developing lesson plans for tutorials. She tolerated boxes of dried specimens and the odor of mothballs until she could stand it no more, and then proposed and supported the conversion of the Tulloss garage to a herbarium with temperature and humidity control to house RET's mycological collections. | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| intro |
Olive text indicates a specimen that has not been
thoroughly examined (for example, for microscopic details) and marks other places in the text
where data is missing or uncertain. The following text is based upon molecular data from Stephen Russell's group (Purdue) and ?? and other original research by R. E. Tulloss. | ||||||||
| pileus | 65 - 70 mm wide, white to pale grayish white to pale gray, campanulate with incurved margin at first becoming planoconvex with decurved margin, somewhat translucent prior to full expansion, subviscid; context very firm, off-white, with pileipellis sometimes dark gray in cross-section, sometimes with watery line immediately above lamellulae, thinning evenly from stipe to margin; margin nonstriate, nonappendiculate; universal veil often absent. | ||||||||
| lamellae | free to receding, with decurrent line on stipe, crowded, pale cream with pinkish tint in mass, white to pale cream in side view (nearly concolorous with stipe context), staining or bruising reaction not observed, ca 5 mm broad, broadest at mid-point, narrowly lozenge-shaped; lamellulae subtruncate to rounded attenulate to subattenuate to attenuate, of diverse lengths, plentiful, with shape and length apparently unrelated. | ||||||||
| stipe | 71 - 80 × 11 - 12 mm, white, subcylindric or narrowing upward, with surface innately fibrous, sometimes with faint pinkish brown stains on lower stipe: context solid, pale cream, bruising or staining not observed, insect damage not observed; bulb elongate or narrowly ellipsoid, with uneven surface, 26 - 35 × 15 - 17 mm; partial veil white, membranous persistent, exact position not recorded (apical, subapical or superior), striate above, with underside bearing fine fibrils near attachment to stipe and otherwise smooth; universal veil white, submembranous-friable, detersile, sometimes leaving faint margin around top of bulb or patches in adjacent substrate or small fragments on bulb surface. | ||||||||
| odor/taste | Odor indistinctly fungoid. Taste not recorded. | ||||||||
| macrochemical tests |
KOH - negative on pileipellis. Paracresol (1%) - faint at first, slightly intensifying over 15 min., center of pileus under pileipellis, bulb and stipe base, isolated spots in stipe apex. Sygingaldazine (0.1%) - negative throughout basidiome. Test vouchers: Tulloss 8-16-81-A, Tulloss 7-14-96-D.. | ||||||||
| lamella trama | bilateral, divergent; ... | ||||||||
| subhymenium | pseudoparenchymatous (cellular); ??. | ||||||||
| lamella edge tissue | sterile. | ||||||||
| basidiospores | RET: [20/1/1] 7.0 - 9.1 (-9.4) × 5.2 - 6.3 (-6.6) µm, (L = 8.0 µm; L’ = 8.0 µm; W = 5.7 µm; W’ = 5.7 µm; Q = (1.19-) 1.25 - 1.55; Q = 1.40; Q’ = 1.40), hyaline, colorless, smooth, thin-walled, amyloid, broadly ellipsoid to ellipsoid, adaxially flattened; apiculus sublateral, cylindric, proportionally small; contents ??; white in deposit. | ||||||||
| ecology | Solitary to scattered. Tlaxcala, Mexico: At 2775 m elev. In dark, moist, friable loam of Pinus-Abies-Quercus forest. Massachusetts, U.S.A.: In mixed forest with Amanita jacksonii. Pennsylvania, U.S.A.: In Tsuga canadensis forest. | ||||||||
| material examined | MEXICO: TLAXCALA—Mpio. Tlaxco - El Rodeo, Cerro El Peñon [19.7000° N/ 98.2211° W, 2775 m], 14.vii.1996 Dr. Arturo Estrada Torres s.n. [Tulloss 7-14-96-D] (TLXM; RET 254-6, nrITS & nrLSU seq'd.). U.S.A.: MASSACHUSETTS—Unkn. Co. - ca. border Berkshire & Franklin Cos., W of Charlemont, Mohawk Trail St. Pk., 16.viii.1981 M. A. King & R. E. Tulloss 8-16-81-A (RET 323-3, nrITS & nrLSU seq'd.); NEMF 1986, 16.viii.1986 foray participant [Tulloss 8-16-86-J] (RET 138-9, nrITS & nrLSU). PENNSYLVANIA—Monroe Co. - Pocono Region [41.1271° N/ 75.4907° W, 514 m], 19.vii.2015 David Wasilewski s.n. [mushroomobserver #210821] (RET 709-6, nrITS-LSU seq'd.). | ||||||||
| discussion |
The semi-translucent appearance of the young cap,
the floral odor of the basidiome, the absence of
yellow in the universal veil in all known
specimens, and the lack of bruising reaction
suggest that this taxon is distinct from
A.
rubescens var. alba Coker and other
rubescent North American taxa.
The following figure provides a sporograph
comparison between the latter taxon and A.
maryaliceae: The lack of bruising on the fruiting body distinguishes the present species from other, recently distinguished white rubescent taxa such as A. ostendemihi and the white-capped variant of A. aureosubucula. The present taxon could be the poorly known A. spissa var. alba Coker with the following exceptions. Coker describes his material as In an additional collection in NCU annotated in Coker’s hand ("North Carolina, ??, deciduous woods SE of the Graded School, 12.vii.1920 H. R. Totten 4386," as “Amanita spissa var. alba”) Coker wrote: “This is exactly like Hartsville no. 11—Cap 7 cm broad, smooth, white with a few light brown stains. No volva patches. Margin faintly striate, surface pitted. Veil ?? + staining exactly as in my 11, even to the faint pinkish flocculence from the volva on the good sized basal bulb. This var. is apparently near but certainly not the same as ?? of no. 4377 which differs in staining much more freely + quickly (gills in var. alba hardly stain at all; caps stain slowly and very little), distinct warts + shorter stem + smaller size. Stem including bulb 13 cm long. Spores white, elliptic, smooth, 4.4 - 6.5 × 6.3 - 9.2 µm [est. L = 7.8 µm; est. W = 5.4 µm; est. Q = 1.42].” [His 4377 might be A. salmonescens Tulloss or even A. canescens Dav. T. Jenkins. Remember that by 1917 he had described A. rubescens var. alba, which can be robust.]
| ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita maryaliceae |
| bottom links | [ Keys & Checklists ] |
| name | Amanita maryaliceae |
| bottom links | [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.