


| name | Amanita huijsmanii |
| name status | nomen acceptum |
| author | F. Massart & Rouzeau |
| english name | "Huijsman's Ringless Amanita" |
| images |
 2. Amanita huijsmanii, southwestern France. |
| cap | The cap of A. huijsmanii is 45 - 60 mm wide, subcampanulate at first, then convex, with a distinct broad umbo in some specimens, dry, with a strongly striate margin (25±% of the radius); the cap is metallic gray, occasionally slightly darker in the disc. The volva is usually absent. |
| gills | The gills are free, not too close, white with a flesh-cream tone in mass in mature specimens, and rounded at the cap margin. The short gills are truncate at a right angle or at a slight slant. |
| stem | The stem is 55 - 70 × 7 - 9 mm, white, narrowing slightly and evenly upward, ornamented with fine white fibrils, and exannulate. The flesh is white and hollow. The volva is membranous, white, and often completely lacking an internal limb. |
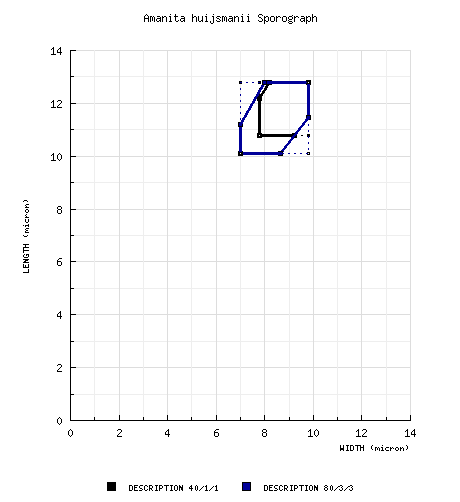
| spores | The spores measure (8.5-) 10.1 - 12.8 (-15.0) × (6.5-) 7.0 - 9.8 (-10.9) µm and are inamyloid and broadly ellipsoid to ellipsoid (infrequently subglobose, infrequently elongate, rarely cylindric). Clamps are rare or absent at bases of the basidia. |
| discussion |
Amanita huijsmanii was originally described from France. It is known from
southern Europe. For comparison, see A. argentea Huijsman, A. mairei Foley, and A. supravolvata Lanne.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita huijsmanii | ||||||||||||||||||||
| author | F. Massart & Rouzeau. 1990 ["1989"]. Bull. Soc. Linn. Bordeaux 17(3): 159, pl. 1. | ||||||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||||||
| english name | "Huijsman's Ringless Amanita" | ||||||||||||||||||||
| synonyms |
≡Amanita argentea f. huijsmanii (F.
Massart & Rouzeau) Contu 1999. 1999. Boll. Acc. Gioen. Sci. Nat. 32: 20. The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||||||||||||||
| etymology | genitive of a Latinized name; hence "Huijsman's" or "of Huijsman" | ||||||||||||||||||||
| MycoBank nos. | 129514 | ||||||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||||||
| holotypes | LIP; isotype (fragments) in RET | ||||||||||||||||||||
| type studies | Tulloss. 1994. Mycotaxon 52: 334, fig. 19. | ||||||||||||||||||||
| revisions | Hanss, J.-M. and P.-A. Moreau. 2020 [2017]. Bull. Soc. Mycol. France 133 (1-2): 67-141. | ||||||||||||||||||||
| selected illustrations | F. Massart. 2000. Boll. Gruppo Micol. G. Bresadola 43(2): 256. | ||||||||||||||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text representing a type study by Tulloss. The same field may also contain black text, which will represent a revision of the species by Tulloss. Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following is based on the cited literature, molecular study of Der. Linas V. Kudzma, and other original research by R. E. Tulloss. | ||||||||||||||||||||
| lamella edge tissue | sterile. | ||||||||||||||||||||
| basidiospores |
Tulloss (1994): [40/1/1] (8.5-) 10.8 - 12.8 (-15.0) × (6.8-) 7.8 - 9.8 (-10.5) μm,
(L = 11.5 μm; W
= 8.7 μm; Q = (1.10-) 1.17 - 1.56 (-1.65); Q = 1.30), hyaline, colorless, thin-walled, smooth, inamyloid, broadly ellipsoid to ellipsoid, occasionally subglobose, infrequently elongate, ocasionally adaxially flattened, occasionally expanded at one end; apiculus sublateral, cylindric; contents guttulate; white in deposit. composite of data from all material revised by RET: [80/3/3] (8.5-) 10.1 - 12.8 (-15.0) × (6.5-) 7.0 - 9.8 (-10.9) μm, (L = 11.3 - 11.5 μm; L' = 11.4 μm; W = 7.8 - 8.7 μm; W' = 8.4 μm; Q = (1.10-) 1.17 - 1.60 (-2.13); Q = 1.30 - 1.46; Q' = 1.36), hyaline, colorless, thin-walled, smooth, inamyloid, broadly ellipsoid to ellipsoid, occasionally subglobose, infrequently elongate, ocasionally adaxially flattened, occasionally expanded at one end; apiculus sublateral, cylindric; contents guttulate; white in deposit. | ||||||||||||||||||||
| ecology | France: On right bank of Garonne R. in Entre-deux-mers, August and September, in deciduous or mixed woods, on clayey-calcareous soil. Portugal: Scattered. At 309 m elev. Portugal: Under Castanea. | ||||||||||||||||||||
| material examined | FRANCE: DORDOGNE—Payzac, 17.x.1997 Christian Rouzeau s.n. [Massart 97058] (in herb. Massart; RET 272-7). GIRONDE—St-Vivien-de-Monségur, 25.viii.1989 Francis Massart 89A1 (holotype, LIP, nrITS seq'd.; isotype, RET 011-1(a), fragments), 89A2 (paratype, RET 011-1(b)), 17.vii.1997 Francis Massart 97043 (in herb. Massart; RET 272-8). PORTUGAL: Serra de Montejunto {39.2035° N/ 9.05° W, 309 m}, 4.x.2014 Zacarias Lepista s.n. [mushroomobserver #181567] (RET 650-10, nrITS & nrLSU seq'd.). | ||||||||||||||||||||
| citations | —R. E. Tulloss | ||||||||||||||||||||
| editors | RET | ||||||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita huijsmanii |
| name status | nomen acceptum |
| author | F. Massart & Rouzeau |
| english name | "Huijsman's Ringless Amanita" |
| images |
2. Amanita huijsmanii, southwestern France. |
| photo |
Zacarias Lepista - (1) Serra de Montejunto,
Portugal.
(RET 650-10). [Note: For untrimmed and
unedited photographs see
here.—ed.] Francis Massart - (2) southwestern France. |
| name | Amanita huijsmanii |
| bottom links | [ Keys & Checklists ] |
| name | Amanita huijsmanii |
| bottom links | [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.