


| name | Amanita aprica |
| name status | nomen acceptum |
| author | J. Lindgr. & Tulloss |
| english name | "Sunshine Amanita" |
| images |
  1. Amanita aprica, note thin layer of volva remaining attached to cap surface, Oregon, U.S.A.  2. Amanita aprica, Oregon, U.S.A.  3. Amanita aprica, Oregon, U.S.A.  4. Amanita aprica, Oregon, U.S.A. |
| intro |
Amanita aprica is a species of old growth forest illustrated and described in (Tulloss and Lindgren, 2005). It is known from SW Canada and the Pacific Northwest of the contiguous U.S. In addition to its striking cap color, its tendency to occur in sunlight along paths and in breaks in forest, note the frost-like layer of volva that often remains attached to the cap surface for a long time. The uppermost part of the cap's skin and the lowermost part of the volval remnants on the cap are actually intergrown. |
| cap | The cap is 50 - 150 mm wide, bright yellow to egg yellow to lemon yellow or bright orangish yellow, occasionally orange (especially in the middle), colors sometimes fading and duller after exposure, globose at first, then convex to plano-convex, finally flat with or without center depressed. The flesh is 6 - 20 mm thick above the stipe and white except immediately below the cap skin where the flesh is yellow. The cap''s margin is not or barely striate. Volval remnants on the pileus appear as a frost-like layer of downy fibrils and/or a covering of confluent warts (white to creamy white to yellowish white). The intimate attachment to the cap skin causes the volval remnants to stretch (becoming thinner as they do so) with the expansion of the cap. |
| gills |
The gills are free to seceding, close to subdistant, white to creamy white in mass and in side view, 5 - 12 mm broad, broadest at midpoint, and bear a fimbriate (sometimes uneven) edge. Short gills are truncate to excavate-truncate and numerous. |
| stem |
The stipe is 33 - 91 × 14 - 35 mm, white to cream to creamy tan, and bruising light tan where handled, usually cylindric. The stipe's bulb is often indistinct at maturity, taking the form of a subclavate base to the stipe. The stipe's context is white to pale yellow and firmly stuffed with white tissue when young; it may become partially or entirely hollow later in development. The annulus is superior to median, white to cream, skirt-like, felted-membranous at first, often collapsing on the stipe, and sometimes evanescent. Volval remnants at the stipe's base usually appear as a low free limb encircling the top of the bulb. The remnants are white to creamy tan to pale tan and are seldom rolled outward. They may also appear as detersile rags on the lower stipe, may be entirely left behind the soil by a collector, or as detersile rings above the free limb mentioned above. |
| odor/taste |
The species is POISONOUS producing nausea, vomiting, intestinal cramps, muscle spasms, and diarrhea. |
| spores |
The spores measure (8.0-) 9.5 - 13.0 (-21) × (5.0-) 6.5 - 8.5 (-12.5) µm and are ellipsoid to elongate (occasionally broadly ellipsoid) and inamyloid. Clamps are infrequent at the bases of basidia and rather difficult to find. |
| discussion |
Amanita aprica occurs mainly with conifers (Douglas Fir and several species of Pine) and is quite common in the Cascade Mtns. of Oregon and Washington states (U.S.A.). It also occurs in the Sierra Mtns. in California. It may be found as a solitary mushroom or growing gregariously at altitudes from 600 - 1850 m elev. It is reported from the spring and very early summer. None of the material collected in the fall and reviewed by the species'' authors proved to belong in A. aprica.
The fact that rings of volval tissue may be left on the lower stipe during development of A. aprica has caused it to be confused with various taxa classed as A. muscaria (in a broad sense). The species in the A. muscaria "group" do not have the cap skin and volva connected as is found in A. aprica and can be distinguished in the field by absence of warts or presence of scattered warts instead of having the distinctive layer of volva typical of A. aprica. Under the microscope, all the taxa in the muscaria "group" have common basidial clamps. Amanita aprica is most similar to taxa of the A. gemmata "group." The only species of this group that has been named in the U.S. is A. russuloides (Peck) Sacc., a species occurring only in eastern North America—as far as is known. The matter is made confusing for North Americans by the fact that as many as three or four other taxa are called "A. gemmata" in their collecting regions. However, none of these mushrooms appears to actually belong to the European taxon. To make matters worse, taxa with a range of different colors are all called gemmata in Europe. The status of these taxa is still not clarified. |
| brief editors | RET |
| name | Amanita aprica | ||||||||||||||||||||||||
| author | J. Lindgr. & Tulloss in Tulloss & J. Lindgr. 2005. Mycotaxon 91: 194 | ||||||||||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||||||||||
| english name | "Sunshine Amanita" | ||||||||||||||||||||||||
| etymology | apricus, growing in the sunshine, exposed to the sun, sunny; because of the species' being found most often in places exposed to direct sun and because of the bright “apricot-like” color of the pileus. | ||||||||||||||||||||||||
| MycoBank nos. | 343980 | ||||||||||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||||||||||
| holotypes | WTU; isotypes in L, NY, & RET | ||||||||||||||||||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. Unless indicated otherwise, this description is drawn directly from the protolog. The original description by Tulloss and Lindgren (2005) is available as a PDF. (open) | ||||||||||||||||||||||||
| pileus | 50 - 150 mm wide, bright yellow to egg yellow to lemon yellow or bright orangish yellow, occasionally orange (especially over disc), colors sometimes fading and becoming duller after exposure to sun, globose at first, then convex to plano-convex, finally plane with or without disc depressed, glabrous, tacky to subviscid where pileipellis exposed at maturity or in moist weather; context white, yellow under pileipellis, 6 - 20 mm thick at stipe, thinning evenly to less than 2 mm at margin; margin nonstriate or faintly striate (even in age), entire, inrolled at first, slightly uplifted in age, sometimes bearing fragments of partial veil; universal veil as thin appressed downy fibrils or small floccose warts or patches or as a nearly complete floccose-felted to “woolly-felted” covering, or as a nearly complete covering of confluent broadly subpyramidal warts, white to creamy white to yellowish white, detersile, quite commonly remaining attached over much of pileipellis and becoming thinner as stretched by expanding pileus. | ||||||||||||||||||||||||
| lamellae | free to seceding, with faint decurrent lines on stipe apex, close to subdistant, white to creamy white in mass and in side view, unchanging when cut or bruised, 5 - 12 mm broad, broadest at midpoint, with fimbriate sometimes uneven edges; lamellulae truncate to excavate-truncate, numerous. | ||||||||||||||||||||||||
| stipe | 33 - 91 × 14 - 35 mm, white to cream to creamy tan, bruising light tan where handled, usually cylindric, occasionally narrowing upward, surface pruinose to flocculose to scurfy below partial veil, flocculence in young specimens easily removed by handling; bulb often not distinct, merely clavate base of stipe, (10-) 16 - 34 × 18 - 40 ( 50) mm, with either rounded or pointed base, sometimes slightly radicating; context white to pale yellow, infrequently with brownish stains in spots on cut surface, firmly stuffed with white tissue when young, becoming hollow or partially hollow; partial veil superior to median, white to cream, felted-membranous at first, soon fragile, skirt-like, sometimes collapsing on stipe, sometimes evanescent; universal veil as low free limb encircling top of bulb, white to cream to creamy tan to pale tan, seldom rolled outward, 1 - 3 mm thick at about half height of limb, 25 - 38 mm from base of bulb to highest point on limb, sometimes also as detersile rings on lower stipe above limb, also often left in soil or as patches or warts on stipe. | ||||||||||||||||||||||||
| odor/taste | Odor and taste not distinctive. | ||||||||||||||||||||||||
| macrochemical tests |
10-20% KOH - partial veil turning orange in less than 5 min., context of pileus and stipe pale yellowish. NH4OH - context of pileus and stipe slightly yellow in 5 min., fading. H2SO4 (dilute) - negative. FeSO4 - negative. Ethanol - negative. Phenol - wine red spot in pileus context and upper stipe after 10 min. Spot test for tyrosinase (paracresol) - positive (reddish brown) in stipe context near stipe base, partial veil, and pileus context in 10 min. Spot test for laccase (syringaldazine) - negative throughout basidiocarp. Macrochemical test voucher: Lindgren 92-5. POISONOUS: producing nausea, vomiting, intestinal cramps, muscle spasms, diarrhea, disorientation, and hallucinations, with these accompanied by drowsiness and the desire to sleep. | ||||||||||||||||||||||||
| pileipellis | 295 - 300 µm thick at approximately midradius, up to 440 ( 535) µm thick in disc, entirely ungelatinized well beyond initiation of sporulation in most specimens and, hence, lacking distinctive supra-/subpellis, with most elements subradially oriented, with occasional regions below upper surface having many elements distinctly nonradial; filamentous, undifferentiated hyphae 1.8 - 7.0 µm wide, branching, densely packed vertically, with many at and near upper surface connected to universal veil well into maturity in most specimens, with hyphae and inflated cells like those of universal veil sometimes arising below uppermost hyphal layers (Fig. 2b); vascular hyphae 3.0 - 19.6 µm wide, branching, sinuous, scattered to locally common, sometimes entangled, occasionally crossing ungelatinized into universal veil well into maturity. | ||||||||||||||||||||||||
| pileus context | filamentous, undifferentiated hyphae 0.6 - 9.3 µm wide, branching rather frequently, plentiful, sometimes in fascicles, sometimes with yellowish subrefractive walls, forming open lattice-like structure; acrophysalides plentiful, terminal and solitary, narrowly clavate to clavate (up to 222± × 51± µm) or broadly clavate to ellipsoid (up to 108 × 47 µm); vascular hyphae 3.0 - 23 µm wide, branching, sinuous, infrequent, locally common (especially in upper part of context). | ||||||||||||||||||||||||
| lamella trama | bilateral, markedly divergent, rather broad; wcs = 95 - 135 µm (very good rehydration); central stratum including numerous intercalary partially inflated to narrowly ventricose cells (e.g., 154 × 18.2 µm); angle of divergence from very shallow to about 45°; filamentous, undifferentiated hyphae 1.0 - 10.5 µm wide, frequently branching, sometimes with yellowish tint, sometimes with constrictions at septa, occasionally markedly changing diameter at septa; divergent inflated cells broadly to narrowly clavate to narrowly ventricose to only slightly inflated, sometimes slightly curved, up to 117 × 32 µm (commonly about half this size), apparently all intercalary; vascular hyphae 4.5 - 9.3 µm wide, rare; clamps infrequent in subhymenial base, not observed elsewhere. | ||||||||||||||||||||||||
| subhymenium | wst-near = 80 - 115 µm (very good rehydration); wst-far = 100 - 135± µm (very good rehydration); (2 ) 3 - 4 (-5) layers of cells thick, with basal layer cells arising from hyphal segments or inflated intercalary cells, generally appearing subcellular to cellular, with (however) basidia arising from both subglobose to barrel-shaped cells and occasionally from only slightly inflated hyphal segments; clamps infrequent to rare. | ||||||||||||||||||||||||
| basidia | 48 - 70 × 10.0 - 11.5 µm, projecting up to 12.0 - 16.0 µm beyond surrounding basidioles, dominantly 4-, occasionally 2-sterigmate; sterigmata up to 5.8 × 4.0 µm; clamps and proliferated clamps unevenly distributed, infrequent, sometimes small and very thin-walled, requiring persistent search. | ||||||||||||||||||||||||
| universal veil | On pileus: with substantial number of filamentous, undifferentiated hyphae and some vascular hyphae connecting to pileipellis often well into maturity, with gelatinization beginning in hyphae near base simultaneously with some hyphae of pileipellis surface, with elements having subvertical to vertical orientation; filamentous, undifferentiated hyphae 3.0 - 8.0 µm wide, branching, plentiful to dominant in base, plentiful above basal region; inflated cells plentiful, terminal singly, broadly to narrowly ellipsoid (up to 78 × 40 µm), broadly to narrowly clavate (up to 105 × 50 µm), sometimes slightly constricted; vascular hyphae 4.2 - 11.5 µm wide, scattered to locally clustered, sinuous, occasionally branching, occasionally entangled locally. On stipe base, exterior surface: filamentous, undifferentiated hyphae in broad (but not very thick) interwoven fas-cicles, partly gelatinized. On stipe base, interior: filamentous undifferentiated hyphae 1.8 - 12.6 µm wide, branching, dominating, sometimes constricted at septa, sometimes with yellowish walls, plentiful, often dominating; inflated cells terminal, singly or in chains of two, globose to broadly globose to pyriform (up to 55 × 52 µm) or broadly ellipsoid to broadly clavate to ellipsoid to elongate (up to 64 × 39 µm), plentiful; vascular hyphae not observed. | ||||||||||||||||||||||||
| stipe context | longitudinally acrophysalidic; filamentous, undifferentiated hyphae 1.4 - 9.1 µm wide, branching, sometimes in narrow fascicles, sometimes with yellowish subrefractive walls, dominantly longitudinally oriented, dominating near surfaces, plentiful in interior; acrophysalides plentiful, smaller and less frequent toward surfaces, plentiful in interior, up to 195 × 37 µm; vascular hyphae 3.2 - 28 µm wide, sinuate to hypersinuate, sometimes loosely coiled (corkscrew-like), unevenly distributed, scattered to locally common, yellowish. | ||||||||||||||||||||||||
| partial veil | filamentous, undifferentiated hyphae 2.1- 4.4 µm wide, branching, dominating, disordered except for occasional fascicles up to 10 hyphae wide, sometimes with yellowish subrefractive walls; inflated cells plentiful, narrowly cavate, terminal, usually singly, occasionally subtended by a smaller somewhat inflated hyphal segment, up to 88 × 17.6 µm wide, infrequently producing hyphae laterally or apically; vascular hyphae not observed. | ||||||||||||||||||||||||
| lamella edge tissue | sterile. | ||||||||||||||||||||||||
| anatomical figures |
, elements of lamella trama and hymenium (some elements displaced by sectioning—for example, apparently sinuate intercalary element in subhymenial base). Scale bar=20 μm.") 1. Amanita aprica (holotype), elements of lamella trama and hymenium (some elements displaced by sectioning—for example, apparently sinuate intercalary element in subhymenial base). Scale bar=20 μm. , elements of lower part of universal veil from pileipellis illustrating interconnection with pileipellis. Scale bar=20 μm.") 2. Amanita aprica (holotype), elements of lower part of universal veil from pileipellis illustrating interconnection with pileipellis. Scale bar=20 μm. , elements of universal veil from pileus (upper portion, crush mount, some hyphae not shown). Scale bar=20 μm.") 3. Amanita aprica (holotype), elements of universal veil from pileus (upper portion, crush mount, some hyphae not shown). Scale bar=20 μm. , elements of universal veil from interior of limb above stipe's bulb (crush mount, some hyphae not shown). Scale bar=20 μm.") 4. Amanita aprica (holotype), elements of universal veil from interior of limb above stipe's bulb (crush mount, some hyphae not shown). Scale bar=20 μm. , elements of partial veil, excluding any cells remaining from former attachment to edges of lamellae (crush mount, some hyphae not shown). Scale bar=20 μm.") 5. Amanita aprica (Trudell 96-165-01), elements of partial veil, excluding any cells remaining from former attachment to edges of lamellae (crush mount, some hyphae not shown). Scale bar=20 μm. | ||||||||||||||||||||||||
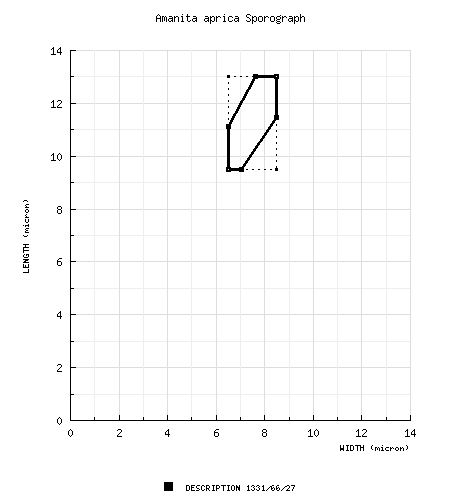
| basidiospores | [1331/66/27] (8.0-) 9.5 - 13.0 (-21) × (5.0-) 6.5 - 8.5 (-12.5) µm, (L = (9.8-) 10.1 - 12.2 (-13.1) µm; L´ = 11.1 µm; W = (6.6-) 6.8 - 8.1 (-8.4) µm; W´ = 7.4 µm; Q = (1.22-) 1.35 - 1.71 (-2.13); Q = (1.38-) 1.42 - 1.62 (-1.80); Q´ = 1.52), hyaline, colorless, smooth, thin-walled, inamyloid, ellipsoid to elongate, occasionally broadly ellipsoid, often adaxially flattened, sometimes swollen at one end, occasionally langeniform, infrequently “giant”; apiculus sublateral, cylindric to truncate-conic, proportionately rather large; contents granular to mono- or multiguttulate, dominantly monoguttulate with additional small granules; white to creamy white in deposit. | ||||||||||||||||||||||||
| ecology | Solitary to gregarious, at 600 - 1850 m elev. Often in sunny locations such as road cuts, along trails, campgrounds, and under other breaks in the forest canopy; occurring with conifers, mainly Pseudotsuga menziesii (Mirb.) Franco, less frequently with Pinus (e.g., P. contorta Dougl. ex Loud., P. monticola Dougl. ex D. Don, and P. ponderosa Dougl. ex Lawp;s.). For example, basidiocarps of Lindgren 90-23 collected under P. monticola with understory of Achlys triphylla (Sm.) DC, Pachistima myrsinites (Pursh) Raf., Vaccinium sp., Pyrola sp., and grass. Quite common in the Cascade Mountains of Oregon and Washington. | ||||||||||||||||||||||||
| material examined |
CANADA:
BRITISH COLUMBIA—Brackendale, 2.v.1993 P.
Kroeger 1658 (paratype, DAVFP n.v.; paratype, RET
327-2; paratype, UBC n.v.).
U.S.A.:
CALIFORNIA—El Dorado Co. - 1.6 km
E of Ice House Reservoir [38.8213° N/ 120.3767° W,
1510 m], 14.v.2013 Mike McCurdy s.n.
[mushroomobserver.org
#134573]
(RET 534-7, nrITS & nrLSU seq'd.). Sierra Co. - N of Downieville, ca.
Hwy. 49, 5.vii.2003 Mark Lockaby “A” (paratype, RET
369-10); Sierra Nevada Field Camp on Hwy. 49,
6.vi.1986 R. E. Halling 4659 (paratype, NY).
Siskiyou Co. - Shasta National
Forest, E of McCloud, ca. 1.6 km from Rte. 89/For.
Rd. 13 jct, E of For. Rd. 13 | ||||||||||||||||||||||||
| discussion |
Amanita aprica is assignable to Amanita section Amanita. For many years in the Pacific Northwest of the U.S.A., A. aprica has been identified as other taxa of that section: Amanita muscaria var. formosa Pers., A. gemmata, or A. jonquillea Quél.—also spelled “junquillea”—a posterior synonym of A. gemmata. It seems very likely to have been treated by Nakamura (1965) as both a “form” of A. junquillea sensu Nakamura having ellipsoid spores and as “form B” of A. muscaria var. formosa sensu Nakamura. Regrettably, none of Nakamura’s personal collections are to be found at WTU. Although its macroscopic appearance sometimes suggests otherwise (and, indeed, some collections thought at first to be A. aprica and examined during the work on this paper proved to be A. muscaria subsp. flavivolvata Singer), A. aprica is readily segregatable from all the described varieties of A. muscaria and phenetically closely related taxa by the fact that these all bear common to plentiful basidial clamps. One collection thought at first to be A. aprica proved to be A. breckonii Thiers & Ammirati (1982). Tulloss has examined the type of A. breckonii and found the latter species differs from A. aprica at least in having The following figure compares sporographs for A. aprica and A. breckonii. The supposed double annulus of A. breckonii comprises a weakly structured partial veil and, below this, a substantial ring of tissue of the limbus internus of the universal veil. The latter has a triangular cross-section having its apex pointing upward—strongly suggesting a second partial veil at first glance. In the several exsiccata of A. breckonii examined by Tulloss, the “lower annulus” is not always present. Amanita gemmata (the European “taxon” may be a complex of several entities) can be distinguished from A. aprica by means of the following characteristics: Kauffman (1926: 122-123) describes material from Mt. Hood determined by him as A. jonquillea with spores 10-12 × 7-8 mm (his comments indicate that he excludes extremes of length and, hence, presumably, width). It is very possible that Kauffman’s material determined as jonquillea is assignable to A. aprica. The habit and spores described for A. muscaria var. formosa sensu Nakamura are rather good matches to the habit and spores of A. aprica. In addition, Nakamura’s “form B” is the only yellow entity of Amanita section Amanita described by Nakamura that occurs in the April-June period—apparently the most common fruiting period for A. aprica—indeed, all our collections from Oregon and Washington were made in this period. While accepting with minimal comment Nakamura’s observations on A. muscaria, Breckon (1968: 51-57) appears to have interpreted A. aprica as “montane specimens” of “Amanita junquillea sensu A. H. Sm.” Among the characters noted for the “montane specimens,” Breckon includes a pileus with nonstriate margin and universal veil tending to cover much of the surface. The fruiting period for these specimens is described as “late spring through the autumn,” which is compatible with the dates of collection of the California paratypes of A. aprica. North American taxa referred to A. gemmata include A. russuloides (Peck) Sacc. (1887) (Jenkins 1977), A. xylinivolva Tulloss, Ovrebo and Halling (1992), and several apparently distinct, undescribed species. Both of the above-named species are smaller and more gracile than A. aprica; both have striate margins; and both have proportionately broader spores. Asian species, including those recently described by Zhu L. Yang and others, do not include any taxa that are comparable with A. aprica in all of the following: color, stature, form of universal veil, and spore dimension and shape. Lindgren 92-5 and 94-08 and Lockaby “A” (so marked by Tulloss) comprise immature material. In correspondence and keys, we have used the code “Amanita sp. NW2” for A. aprica. In several sections of Amanita, there have been noted taxa that characteristically fail to produce, or delay in producing, a gelatinizing zone allowing separation of pileipellis and universal veil. In subsection Vittadiniae Bas of Amanita section Lepidella, as well as in other notable species of that section such as A. rhoadsii (Murrill) Murrill and A. magniverrucata Thiers & Ammirati, the universal veil remains long attached to the pileus context and a pileipellis is reduced to a layer of context including rather densely placed hyphae or, quite often, is entirely absent. In other sections, a freshly opened pileus may have a frosty appearance caused by the dense distribution of hyphal stubs pro-jecting upward from the ungelatinized or minimally gelatinized pileipellis after a submembranous or membranous universal veil has been pulled away during expansion of the basidiome. In section Amanita, in particular, there are a number of taxa that retain much of the universal veil on the pileus because of delayed or absent gelatinization—as has been noted in A. aprica. Some examples are A. basiana Tulloss & M. Traverso, A. farinosa Schwein., A. friabilis (Karst.) Bas, A. nehuta G. S. Ridl., and A. xerocybe Bas. The long-term attachment of pileipellis and universal veil does not appear to be a character that should be considered justification for grouping A. aprica taxonomically with these other taxa. The above group is characterized by the following: Significantly smaller basidiomes (and consequent markedly striate pileus margins), frequent absence of a partial veil (in the single exception, A. basiana, the partial veil is very weakly structured and detersile), powdery universal veil, and spores with Q between 1.0 and 1.35 (95% of A. aprica spores have Q falling above this range). Observations on toxicity: We know of one poisoning involving the present species. On May 30, 1990, Lindgren was involved as the mushroom identifier in a case involving an 80 year-old Bulgarian immigrant who spoke no English. He had eaten a frying pan full of A. aprica spread over both breakfast and lunch and, possibly, sampled the mushroom the previous day. The number of fruiting bodies ingested was not established. There were four remaining mushrooms cleaned, but uncooked upon which the determination was based (Lindgren 90-13). | ||||||||||||||||||||||||
| citations | The original description by Tulloss and Lindgren (2005) is available as a PDF. (open)—R. E. Tulloss | ||||||||||||||||||||||||
| editors | RET | ||||||||||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita aprica |
| bottom links |
[ Keys and Checklists ] [ Pacific coastal states (USA) & region key ] |
| name | Amanita aprica |
| bottom links |
[ Keys and Checklists ] [ Pacific coastal states (USA) & region key ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.