


| name | Amanita pseudoporphyria |
| name status | nomen acceptum |
| author | Hongo |
| english name | "Hongo's False Death Cap" |
| synonyms |
?=Amanita indica R. P. Bhatt, Locq. & T. N. Lakh. |
| images |
 1. Amanita pseudoporphyria, Hunan Prov, China. |
| cap |
Fruiting bodies of Amanita pseudoporphyria are medium-sized to large, sometimes very large. The cap is 50 - 150 mm wide, convex to applanate, greyish to grey to grey-brown, darker in the center, innately fibrillose, and glabrous or covered with a few white to dirty white, membranous volval remnants; the cap's margin is smooth and non-appendiculate, but often with annulus remnants hanging on the margin; the cap's flesh is white. |
| gills |
The gills are free to subfree and white; the short gills are attenuate. |
| stem |
The stem is 80 - 130 × 5 - 20 (-40) mm, subcylindric to attenuate upwards, with a surface that is white and covered with white fibrillose to farinose squamules; the flesh is white; the stem's basal bulb is 10 - 30 (-50) mm wide, ventricose to fusiform, and often somewhat rooting. On the stem's base, the volva is limbate, with a free limb up to 50 cm high. The volva's outer surface is white to dirty white, occasionally greyish, while the inner surface is white. The annulus is apical to subapical, white, fragile, and fugacious or persistent. |
| odor/taste | We have never seen a description of the odor or taste of this mushroom. It is a comestible, market species within its range. Even within that range, it can be confused with toxic (even deadly) species. |
| spores |
Basidiospores (6.0-) 7.0 - 9.0 (-10.5) × (4.5-) 5.0 - 6.5 (-7.5) µm, broadly ellipsoid to ellipsoid, amyloid. Clamps are absent from the bases of basidia. |
| discussion |
This species was originally described from Japan. It is very common and widely distributed in China and often sold, together with A. manginiana sensu W. F. Chiu, in free markets. Amanita pseudoporphyria is very similar to A. manginiana sensu W. F. Chiu, but differs from the latter mainly by its broad ellipsoid to ellipsoid spores and more abundant inflated cells in the volva. Amanita indica R. P. Bhatt, Locq. & T. N. Lakh. may be synonymous with A. pseudoporphyria.—Zhu L. Yang This species is now known from northern India, Nepal, and Thailand in addition to southern China and Japan.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita pseudoporphyria | ||||||||
| author | Hongo. 1957. J. Jap. Bot. 32: 141, fig. 2(a-c). | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Hongo's False Death Cap" | ||||||||
| synonyms |
?=Amanita indica R. P. Bhatt, Locq. & T. N. Lakh. in A. Kumar et al 1990. Amanitaceae India: 71, fig. 9(A-D). The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| MycoBank nos. | 292458, 360945 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes |
Amanita indica—HPUB. Amanita pseudoporphyria—TNS per (Doi. 1991. Bull. Natl. Sci. Mus., Tokyo, Ser. B 17(2): 50]. | ||||||||
| revisions |
Z. L. Yang. 1997. Biblioth. Mycol. 170: 168, figs. 138-142. Z. L. Yang and Y. Doi. (1999). Bull. Natl. Sci. Mus. Tokyo B 25(3): 125-126, fig. 27. Tulloss, here | ||||||||
| selected illustrations |
Imazeki and Hongo. 1965. Color. Illus. Mushr. Japan 2: 40, pl. 12 (fig. 74). Imazeki and Hongo. 1987. Color. Illus. Mushr. Japan 1: 125, pl. 30 (fig. 215). | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material not directly from the protolog of the present taxon and not cited as the work of Dr. Z. L. Yang or another researcher is based upon original research by R. E. Tulloss. NOTE: Spore measurements from papers by Z. L. Yang use his "Times New Roman" face for "Q" and "Q'"—respectively, " | ||||||||
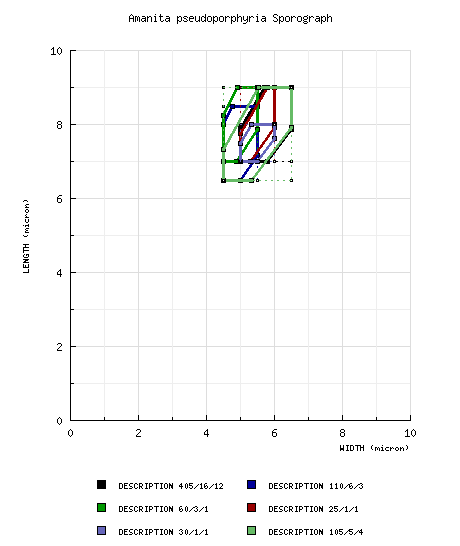
| basidiospores |
from Yang (1997): [405/16/12] (6.0-) 7.0 - 9.0 (-10.5) × (4.5-) 5.0 - 6.5 (-7.5) μm, ( from Yang and Doi (1999) including holotype: [110/6/3] (6.0-) 6.5 - 8.5 (-11.5) × (4.0-) 4.5 - 5.5 (-6.5) μm, ( from Yang and Doi (1999): [60/3/1] 7.0 - 9.0 (-11.0) × (4.0-) 4.5 - 5.5 (-6.5) μm, ( from Yang (2000): [25/1/1] 7.0 - 9.0 × 5.0 - 6.0 μm, ( from Yang et al. (2001): [30/1/1] 7.0 - 8.0 × 5.0 - 6.0 (-6.5) μm, ( from material of India, Nepal, and Thailand (RET): [105/5/4] (5.0-) 6.5 - 9.0 (-10.0) × (3.8-) 4.5 - 6.5 - (-7.2) μm, (L = 7.4 - 8.1 μm; L' = 7.6 μm; W = 5.2 - 5.6 μm; W' = 5.4 μm; Q = (1.19-) 1.22 - 1.63 (-1.80); Q = 1.33 - 1.51; Q' = 1.40), hyaline, colorless to yellowish, smooth, thin-walled, amyloid to strongly amyloid, often adaxially flattened, sometimes swollen at one end; apiculus sublateral, cylindric; contents mono- to multiguttulate, with additional small granules. [NOTE: The extension of spore measurements into a smaller size range in the data of RET, is (at least in part) due to the fact that some of the material he revised was poorly dried and/or poorly preserved.—ed.] | ||||||||
| ecology | Solitary or gregarious. China: At 1800 - 2800 m elev. In conifer and mixed forests with Pinus and Quercus spp. India: ??. Japan: On ground in forests (e.g., Pinus). Nepal: ??. Thailand: ??. | ||||||||
| material examined |
from protolog: JAPAN: HONSHU—Shiga-ken - Ôtsu-shi, Ishiyama-Hiratsu-chô, 23.vii.1956 T. Hongo 1506 (holotype, in herb. Hongo => TNS F-237281). from Yang (1997): CHINA: GUIZHOU—Guiyang (prefecture level) City - Huaxi Distr., unkn. loc., from Yang and Doi (1999): JAPAN: HONSHU—Shiga-ken - Ôtsu-shi, Ishiyama-Hiratsu-chô, 23.vii.1956 T. Hongo 1506 (holotype, in herb. Hongo => TNS F-237281). Tokyo Metropolis (Pref.) - Oume-shi, Kurosawa 3-Chôme, right-side ridge of Kurosawa-gawa R., 2.viii.1997 Y. Doi & K. Miyazaki s.n. (TNS F-182564), Oume-shi, Kurosawa 3-Chôme, left-side ridge of Kurosawa-gawa R., along hiking course, 16.vii.1997 Y. Doi & K. Miyazaki s.n. (TNS F-182479). from Yang (2000): CHINA: JIANGSU—Nanjing (sub-provincial) City - former city of Nanjing, unkn. loc., 28.vi.1936 H. N. Shen 303 p.p. [smaller basidiome] (formerly "IBN 5485"; BPI 750771, as "A. phalloides"). from Yang et al. (2001): CHINA: HAINAN—Lingshui Li Autonomous Co. - Diaoluoshan, 26.v.1988 T. H. Li s.n. (HMIGD 14029, as "Amanita phalloides" in Bi et al. (1997)). RET: INDIA: UTTARAKHAND—Garhwal - Pauri Distr., Khirsu, 28.ix.1993 V. K. & R. P. Bhatt s.n. (GUH M-20189; RET 304-8). NEPAL: CENTRAL REGION—Bagmati Zone, Bhaktapur District, 10 km NE of Bhaktapur City, E of Kathmandu, Nala, 30.vii.1989 H. R. Bhandary s.n. (NHMTU; RET 355-7). THAILAND: CHIANG MAI PROV.—ca. Hang Dong, 20.viii.1995 David Arora s.n. (SFSU; RET 178-10). SAKON NAKHON PROV.—Poo Pan, viii.1998 D. Arora 98-43 (SFSU; RET 351-2). | ||||||||
| discussion |
t.b.d. The Japanese name for this species is "ko-tengutake-modoki" (Doi 1991). | ||||||||
| citations | Translation from German by RET.—Z. L. Yang and R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.