


| name | Amanita virginiana |
| name status | nomen acceptum |
| author | (Murrill) Murrill |
| english name | "Virginian Little Caesar" |
| images |
 1. Amanita virginiana, Washington Crossing St. Pk., Mercer Co., New Jersey, U.S.A. ") 2. Amanita virginiana, Hunterdon Co., New Jersey, U.S.A. (RET 408-4) ") 3. Amanita virginiana, Hunterdon Co., New Jersey, U.S.A. (RET 408-4)   4. Amanita virginiana, North Carolina, U.S.A. |
| intro |
Amanita virginiana is often so small that it can be found in good condition in lawns around mature oaks after the lawn has been mowed. |
| cap |
Its 7 - 50 mm wide cap is dark brown at first and takes on grayer tones in expanding; its margin is striate from one-third to one-half of its radius and more pallid than its center. |
| gills |
The gills are free or nearly so, rather close, white to off-white; and short gills are truncate to subtruncate. |
| stem |
The bulbless stem is 8 - 76 × 2 - 7 mm with a small white volval sac (up to 14 mm high) at its base that some times looks like a small exploded piece of popcorn and with a fragile tiny annulus that is very often lost at maturity. |
| odor/taste | The odor is absent or mild, and the taste is not distinctive. |
| spores |
The spores measure (9.1-) 11.5 - 15.0 (-20.5) × (6.5-) 8.5 - 11.2 (-15.5) µm and are broadly ellipsoid to ellipsoid (rarely subglobose or elongate) and inamyloid. Clamps are common at bases of basidia. |
| discussion |
This is species is one of the small amanitas that has rather plentiful 2-spored basidia and spores that are quite large for the genus. Amanita pachysperma G. F. Atk. and A. subvirginiana (Murrill) Murrill appear to be closely related taxa. One should also compare A. ristichii Tulloss. Amanita virginiana is associated with oak (Quercus) in the eastern U.S.A. It is often plentiful locally, but apparently is seldom (knowingly) collected.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita virginiana | ||||||||||||||||||||||||
| author | (Murrill) Murrill. 1914b. [26 Sept.]. Mycologia 6: 269. | ||||||||||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||||||||||
| english name | "Virginian Little Caesar" | ||||||||||||||||||||||||
| synonyms |
≡Venenarius virginianus Murrill. 1914a [28 July]. N. Am. Flora 10(1): 71.
≡Amanita virginiana (Murrill) Sacc. & Trotter. 1925. Syll. Fung. 23: 4. [Superfluous combination.] The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||||||||||||||||||
| etymology |
Virginia + -ana, suffix indicating possession; hence, "of Virginia" Honoring the Commonwealth of Virginia. | ||||||||||||||||||||||||
| MycoBank nos. | 284084 | ||||||||||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||||||||||
| holotypes | NY; isotype, NCU | ||||||||||||||||||||||||
| type studies | Jenkins. 1979. Mycotaxon 10: 197. | ||||||||||||||||||||||||
| revisions | Tulloss. 1993. Mycotaxon 49: 467, figs. 11-14. | ||||||||||||||||||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material not directly from the protolog of the present taxon and not cited as the work of Dr. Z. L. Yang or another researcher is based on original research by R. E. Tulloss. The information on this page is the result of personal research by R. E. Tulloss. A great deal of the information appeared previously in (Tulloss 1993). | ||||||||||||||||||||||||
| pileus | 7 - 50 mm wide, fuliginous to dark brown to gray-brown to brownish gray (e.g. 10YR 6/2) or umbrinous gray over disk, the gray tones becoming apparent upon expansion, at times with brown tints becoming stronger after collection, pallid to white to sordid cream to tan toward margin, hemispheric at first, then convex to plane, often depressed over disk in all stages of expansion, glabrous, moist to slightly sticky to viscid, becoming shiny when dry; context white, infrequently with unevenly distributed pale gray regions, sordid or brownish under pileipellis, infrequently brownish in disk near apex of stipe, unchanging when cut or bruised, 0.5 - 2.5 mm thick at stipe, thinning evenly for about one-half to two-thirds of radius, then a membrane to margin; margin striate to tuberculate-striate, with long striations [(0.30-) 0.45 - 0.55R], nonappendiculate, decurved at first, flaring upward in maturity; universal veil absent or as pallid, detersile warts or small patches. | ||||||||||||||||||||||||
| lamellae | free to narrowly adnate to adnate, with or without decurrent line on stipe, subdistant to close, white to off-white in mass and in side view, becoming sordid in age, unchanging when cut or bruised, 1 - 5 mm broad, margins fimbriate; lamellulae truncate to rounded truncate to subtruncate, of diverse lengths, originating both at margin and stipe. | ||||||||||||||||||||||||
| stipe | 8 - 76 × 2 - 7 mm, white, faintly browning from handling, cylindric or (infrequently) narrowing downward, slightly expanded at base where confluent with universal veil, sometimes slightly flaring at apex, finely pulverulent near apex, smooth to satiny below, becoming finely fibrillose below (lens), longitudinally striatulate (lens); context white to off-white, unchanging when cut or bruised, hollow or firmly stuffed with white material in 0.5 - 4 mm wide central cylinder, concolorous in larva tunnels; partial veil subapical to superior to median to subinferior, white, usually persistent, thin, small, at first membranous and skirt-like, collapsing with age, occsasionally only present as fibrous-fibrillose line, not apparent on some exsiccata (including some in holotype); universal veil as conspicuous cup or small sac, sheathing or with flaring limbs suggesting a kernel of popped corn, leathery and tough, white, adnate to stipe base, up to 14 mm from stipe base to highest point of volval limb. | ||||||||||||||||||||||||
| odor/taste | Odor absent or "mild" (Began 388). Taste not distinctive. Edibility unknown. | ||||||||||||||||||||||||
| macrochemical tests |
Spot test for tyrosinase (paracresol) - slowly positive (after 5 min. or more) in universal veil, on the stipe surface, pileipellis, and in top and bottom of stipe context; accompanied by faint positive reactions in lamellae and elsewhere. Spot test for laccase (syringaldazine) - negative throughout basidiome. Test voucher: ??. | ||||||||||||||||||||||||
| pileipellis |
20± µm thick, orange-brown in 3% KOH; filamentous, undifferentiated hyphae 1.8 - 8.5 µm wide, tightly interwoven, predominantly subradially arranged, extensively gelatinizing; vascular hyphae not observed. Requires checking to see if supra- and subpellis are differentiated. | ||||||||||||||||||||||||
| pileus context | filamentous, undifferentiated hyphae 1.2 - 7.2 µm wide, plentiful, branching, often in fascicles; acrophysalides dominating, thin-walled, clavate to broadly clavate to ellipsoid, up to 84 × 52 µm; vascular hyphae 4.8 - 5.0 µm wide, rather scarce. | ||||||||||||||||||||||||
| lamella trama | bilateral, difficult to rehydrate in holotype, with wct = 95 - 105 µm in recently collected and well-dried material, with wcs = 30 - 55 µm; divergent elements leaving central stratum at shallow angle, divergent hyphae (almost immediately beginning to branch to form the subhymenium) more easily/frequently seen than divergent, terminal, inflated cells; filamentous, undifferentiated hyphae 1.0 - 3.4 µm wide, with some intercalary segments inflated up to 14.5 µm wide; divergent, terminal, inflated cells up to 43 × 23 µm, ellipsoid to clavate, thin-walled; vascular hyphae 2.1 - 3.2 µm wide, uncommon to very scarce. | ||||||||||||||||||||||||
| subhymenium | [1993 desription seems doubtful to RET, must be redone.] | ||||||||||||||||||||||||
| basidia | 33 - 53 × 9.8 - 14.0 (-14.5) µm, thin-walled, the great majority 2-sterigmate, occasionally 1-sterigmate; clamps common, prominent. | ||||||||||||||||||||||||
| universal veil | On stipe base, external surface: filamentous, undifferentiated hyphae 2.0 - 6.0 µm wide, branching, in fascicles that are sufficiently widely spaced so that the interior structure is visible between them, sublongitudinally oriented, partially gelatinized; clamps present. On stipe base, interior: filamentous, undifferentiated hyphae 2.0 - 7.2 µm wide, collected in fascicles surrounding inflated cells, with walls slightly thickened in those of largest diameter; inflated cells subpyriform to broadly clavate to clavate to narrowly clavate or elongate, dominant, up to 132 × 42 µm, with cell walls thin or slightly thickened (up to 0.5±: µm thick), giving impression of longitudinal orientation to the tissue (however, many cells are not longitudinally aligned); vascular hyphae 2.5±: µm wide, scarce; clamps present. On stipe base, inner surface: like the interior except somewhat collapsed and gelatinized. On pileus: not present in holotype; in other collections, similar to material at stipe base. | ||||||||||||||||||||||||
| stipe context | longitudinally acrophysalidic; filamentous, undifferentiated hyphae 1.8 - 8.0 µm wide, branching, dominant; acrophysalides thin-walled or with slightly thickened walls (up to 0.5 µm thick), plentiful, up to 195 × 55 µm; vascular hyphae 3.5 - 18.0 (-19.0) µm wide, branching, locally rather common; clamps common. | ||||||||||||||||||||||||
| partial veil | not located in holotype [description based on Halling 3489 and Tulloss 7-5-84-C]; upper surface pocked [probably by loss due to gelatinization of ovoid to ellipsoid to clavate cells (e.g., 75 × 45 µm) that permit separation of lamellae from partial veil]; filamentous, undifferentiated hyphae 2.0 - 11.2 µm wide, partially gelatinized, interwoven, frequently branching, subradially arranged at surface, criss-crossed below surface, often in fascicles, rather easily dissociated (at least when not part of a fascicle); inflated cells (other than surface cells) ellipsoid to ovoid to clavate (up to 75 ×45 µm) and narrowly clavate to elongate (up to 305 × 35 µm, but mostly less than 200 × 35 µm), in chains or singly, terminal, thin-walled, partially gelatinizing, very fragile and difficult to detect in exsiccata; vascular hyphae up to 2.5 - 6.3 µm wide, branching, locally common and in tangles and knots; clamps plentiful. | ||||||||||||||||||||||||
| lamella edge tissue | not described. | ||||||||||||||||||||||||
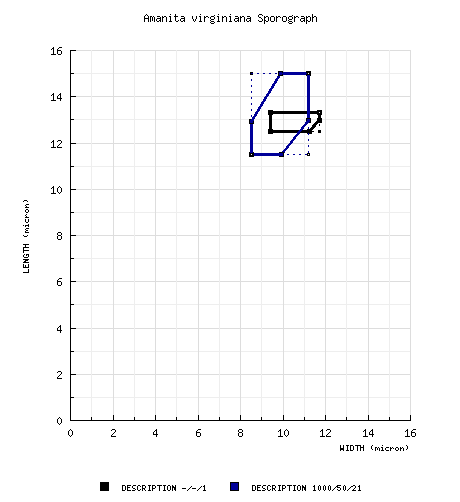
| basidiospores |
from type study of Jenkins
(1979):
[-/-/1] 12.5 - 13.3 (-14.1) ×
9.4 - 11.7 (-13.3) μm, (Q = 1.11 - 1.45;
Q' = 1.28), hyaline, thin-walled,
nonamyloid, subglobose to ellipsoid, often
adaxially flattened; apiculus sublateral,
cylindric; contents guttulate; color in
deposit not recorded. composite data from all material revised by RET: [980/49/20] (9.1-) 11.5 - 15.0 (-20.5) × (6.5-) 8.5 - 11.2 (-15.5) µm, (L = (11.7-) 11.8 - 14.0 (-14.1) µm; L’ = 13.2 µm; W = (9.0-) 9.1 - 10.6 (-10.9) µm; W’ = 9.9 µm; Q = (1.07-) 1.16 - 1.52 (-1.89); Q = (1.16-) 1.17 - 1.44 (-1.55); Q’ = 1.33), hyaline, thin-walled, smooth, amyloid, broadly ellipsoid to ellipsoid, often somewhat adaxially flattened, often expanded at one end; apiculus subapical to sublateral, small to moderately broad, cylindric; contents dominantly monoguttulate, also multiguttulate to granular; white in deposit. | ||||||||||||||||||||||||
| ecology | Scattered to subgregarious to gregarious; up to 1465 m elev. Tennessee and North Carolina northward to eastern Massachusetts and westward to Illinois; late June to late August. Connecticut: in lawn near Quercus. Illinois: Scattered on canyon floor in sandy soil under Fagus grandifolia Ehrh. Massachusetts: In sandy soil under Quercus, Acer, Pinus, and Tsuga. New Jersey: In well-groomed lawn under and around an old, solitary specimen of Q. rubra with Cornus florida and Aesculus hippocastanum nearby. North Carolina: In sandy bank in Pinus woods. Texas: Under Q. nigra and P. taeda. Virginia: In moist, gravelly bank in Quercus woods. | ||||||||||||||||||||||||
| material examined |
from type study of Jenkins
(1979):
U. S. A.:
VIRGINIA— Giles Co. - Mountain Lk.,
8-14.vii.1909 W. A. Murrill 28 (holotype,
NY). RET: U.S.A.: CONNECTICUT—New London Co. - Colchester, Day Pond St. Pk. [41°33'25" N/ 72°25'06" W, 134 m], 24.viii.2007 Paula de Santos s.n. [RET 8-24-07-N] (RET 439-1, nrLSU seq'd.). Unkn. Co. - NEMF 2000, Walk 14, 12.viii.2000 George Davis s.n. [Tulloss 8-12-00-A] (RET 316-1). ILLINOIS—Jackson Co. - Shawnee Nat. For., Little Grand Canyon, ca. 8 km S of Murphysboro, 20.viii.1986 Timothy J. Began 388 (SIU). MASSACHUSETTS—Middlesex Co. - Lincoln, 1± km W of Walden Pond, Old Concord Rd., Adams Woods, 1.viii.1982 Roy E. Halling 3489 (NY). NEW JERSEY—Hunterdon Co. - Oakmoss Mycol. Pres., Lebanon [40°38'50.07" N/ 74°47'50.92" W], 18.vii.2007 R. Ballsley s.n. (RET 408-4). Mercer Co. - Washington Crossing St. Pk., arboretum area [40°18’00”N/74°52’03”W, 32 m], 1.vii.1984 Mary A. King & David C., Mark H., & R. E. Tulloss 7-1-84-E (F; L; NY; RET 021-7; XAL), 5.vii.1984 R. E. Tulloss 7-5-84-A (RET 023-7), -B (RET 056-8), -C (RET 021-5), -D (RET 058-2), -E (RET 021-8), -F (RET 056-6), 23.vi.1985 D. C. & R. E. Tulloss 6-23-85-C (RET 056-3), 5.vii.1992 M. A. King & David C., Sarah E. K. & R. E. Tulloss 7-5-92-B (RET 061-3). Morris Co. - Bernardsville, 23.vii.1984 R. Miller & Dorothy Smullen s.n. [Tulloss 7-23-84-RMDS4] (RET 059-7); Mendham, Meadowood Twp. Pk. [40°47'31" N/ 74°38'43" W, 214 m], 22.vii.1984 NJMA member s.n. [Tulloss 7-22-84-H] (RET 050-2). NEW YORK—Dutchess Co. - Pine Plains, Thompson Pond Preserve, Mt. Stissing [41°57’52” N/ 73°40’57” W, ca. 140-410 m elev.], 2.viii.1990 William Bakaitis 90-67 (RET 047-8). Rockland Co. - Harriman St. Pk., Lake Sebago, 23.vii.1963 Bill Isaacs 2154 (MICH). NORTH CAROLINA— Macon Co. - Highlands, 24.vii.1956 L. R. Hesler s.n. (TENN). SOUTH CAROLINA—Macon Co. - Cliffside Lk. Rec. Area [35°40'43.62" N/ 83°14;12.25" W, 1036 m], 6.viii.2014 L. Gallagher, D. Newman & M. Voller s.n. [mushroomobserver.org #173720] (RET 645-10). TENNESSEE—Sevier Co. - ca. Gatlinburg, GSMNP, Alum Cave Tr. [35°37´48˝ N/ 83°27´04˝ W], 13.vii.2004 David P. Lewis 7-13-04-G (RET 374-8, nrITS, nrLSU & rpb2 seq'd.). TEXAS—Orange Co. - Vidor, 455 Virginia Lane, 20.ix.1987 David P. Lewis 4079 (RET 461-1). VIRGINIA—Giles Co. - ca. Mountain Lk., 8-14.vii.1909 W. A. Murrill 28 (holotype, NY; isotype, NCU). | ||||||||||||||||||||||||
| discussion |
Amanita virginiana is a small to very small mushroom with brownish gray to dark brown to gray-brown to fuliginous disc and pallid pileus margin. Its white stipe is decorated by a superior to median to subinferior, white partial veil that is usually persistent. The universal veil is found as a sheathing volva or as warts on the pileus and, in the latter case, a set of small splayed limbs on the stipe base suggesting a kernel of popped corn. The species appears to be associated with Quercus and Fagus. Inflation of the short hyphal segments of the subhymenium was not seen in all specimens and was usually seen only locally in those specimens in which it occurred. I have the impression that inflation of such cells is more common in A. pachysperma. For further comparison of A. virginiana to the other similar taxa, see the discussion sections following descriptions of A. pachysperma, A. ristichii, and A. subvirginiana. Coker (1917) equated A. spreta var. parva with A. virginiana [for which he reports the spore measurements of 11 - 13 × 7.5 - 10.5 µm (est. Q’ = 1.36)]. Coker determined as A. spreta var. parva his collection Coker 1624 (NCU? n.v.) for which he gives the spore dimensions as 10 - 12.6 × 6.7 - 8.2 µm (Q’ = 1.53). These spores are considerably narrower than those of A. virginiana. The spores of A. spreta var. parva as described by Beardslee are very nearly the size given by Coker [10 - 12 × 7 - 8 µm (est. Q’ = 1.47)]—smaller than those measured for any of the other taxa cited above. As a consequence, it seems unwise to accept Coker’s proposal of synonymy of A. spreta var. parva with A. virginiana without further evidence. Coker 1624 could not be located in NCU. A packet in NCU marked "Amanita spreta var. parva ex type" proved to contain a small box marked in black ink "Venenarius virginianus Part of type" in what appears to be W. A. Murrill’s hand; the material inside was indeed A. virginiana. Coker’s description of A. spreta var. parva reads "This is an extremely little form of A. spreta and is a small plant with a very pale cap, and veil not smoky. Cap 3.5 cm. broad, pale buff in center, fading to nearly white on margin which is strongly striate. Gills attached, about 3 mm. deep, white. Stem 13 cm. long, 6 mm. thick, finely fibrillose below the veil, minutely granular above; a distinct central cylinder that is lightly stuffed. Veil a small collapsed ring 2 cm. from the cap, white. Volva long, narrow, attached to the very base of the stem, which is not bulbous. Spores elliptic, 6.7-8.2 × 10-12.6 µ." Note that the stipe length does not match that of any of the small North American species with numerous bisterigmate basidia, and the cap color is more like that of A. subvirginiana than that of A. virginiana. I could find no evidence that a type of A. spreta var. parva exists or has ever existed; and I could find no authentic material of the taxon. There is no authentic material of A. spreta var. parva in MICH (in which Beardslee’s herbarium is now included). In Beardslee’s files (preserved at MICH), there is no file for A. spreta var. parva; nor is mention of this taxon made in Beardslee’s file on A. spreta (Peck) Sacc. (examined by me). Moreover, it seems very unlikely that, if a type of A. spreta var. parva were known to Coker, he would have designated what he considered to be the type of a synonym (A. virginiana) as the type of A. spreta var. parva in his herbarium. Since the Beardslee taxon is poorly described and apparently lacks a type, it must be considered to be of questionable taxonomic value at present. It is listed under nomina dubia in (Tulloss 1993). If it can be defined clearly in the future and, moreover, is then establsihed as a variety of A. spreta, its basidia would be expected to be dominantly 4-sterigmate. In NY, I found a packet (W. A. Murrill 179) with label indicating that it was a topotype of V. virginianus collected during the same period as was the holotype. Murrill must never have checked the spores of this collection, which is clearly not conspecific with the holotype. As noted by L. R. Hesler (annotation slip attached to packet of Murrill 179), the spores of the single specimen in the packet are globose to subglobose. The Washington Crossing St. Pk. (New Jersey) collections were made in a lawn that was frequently mowed. The specimens were short enough to survive the mowings intact. Since seven collections were made at this site in a period of five days (1-5 July 1984) and since it is possible that they represent fruiting of a single mycelium, it is of interest to see the amount of variation in spore size and shape measured on the specimens of these collections: [380/19/7] (9.1-) 11.5 - 14.5 (-16.2) × (6.5-) 8.8 - 11.5 (-13.0) µm, (L = (12.3-) 12.5 - 13.6 (-13.7) µm; L’ = 13.0 µm; W = (9.5-) 9.6 - 10.7 (-10.9) µm; W’ = 10.1 µm; Q = (1.07-) 1.12 - 1.48 (-1.76); Q = (1.16-) 1.19 - 1.38 (-1.39); Q’ = 1.30). An isolate from Began 388 provided the basis for cultural studies reported in (Began, 1988). | ||||||||||||||||||||||||
| citations | —R. E. Tulloss | ||||||||||||||||||||||||
| editors | RET | ||||||||||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita virginiana |
| bottom links |
[ Keys & Checklists ] |
| name | Amanita virginiana |
| bottom links |
[ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.