


| name | Amanita sp-36 |
| name status | cryptonomen temporarium |
| author | Tulloss |
| images |
") 1. Amanita sp-36, Hopewell Twp., Mercer Co., NJ, U.S.A. (RET 138-4)  2. Amanita sp-36?, but note distinct shoulder on bulb, northeastern U.S.A. ") 3. Amanita sp-36, Middlesex Co., Connecticut, U.S.A. (RET 489-3) |
| intro | Amanita sp-36 is a small to medium-sized species known from the eastern U.S. |
| cap | The dull orange-brown cap is 28-59 mm wide and is hemispheric at first, with a straight or slightly downward curving edge. The cap is tacky at first and becomes dry. The cap’s flesh is yellow directly below the cap’s skin, and white everywhere else. The flesh doesn’t stain when bruised or cut, and is 1-4 mm thick over the stem. The cap’s flesh thins evenly to its edge. The cap’s edge is not grooved, and sometimes has fragments of the ring attached to it. Volval material is present as small orange to yellow warts that may merge together and are easily removed from the cap. Color change in the warts was not observed. |
| gills | The gills of A. sp-36 are very narrowly attached, and there are no downward curving lines on the top of the stem. The gills are close to crowded and 4 - 5.5 mm broad. Short gills end abruptly, are unevenly distributed, are of diverse lengths, and have a gradual tooth projecting along the underside of the cap’s flesh. |
| stem | The white to yellow stem measures 47 - 84 × 4 - 9.5 mm. Yellow pigmentation on the stem can be unevenly distributed or limited to the top or bottom of the stem. The stem is unchanging when cut or bruised. The stem narrows toward its top and is longitudinally lined. The bulb is nearly globose and 15 × 14 mm, or is present merely as a club shaped base to the stem. The stem's flesh is white and is stuffed with cottony material. The thin white to yellow ring can be located from the top of the stem to below the middle of the stem. The ring can become grey-brown with age, it is not lined on top and is fluffy or fibrous below. The volval remnants are present as small yellow granules or flecks on the stem and as loose easily removed patches on and in the soil surrounding the stem’s base. |
| odor/taste | The odor of this mushroom is faintly earthy or undetectable. Its taste has not been recorded. |
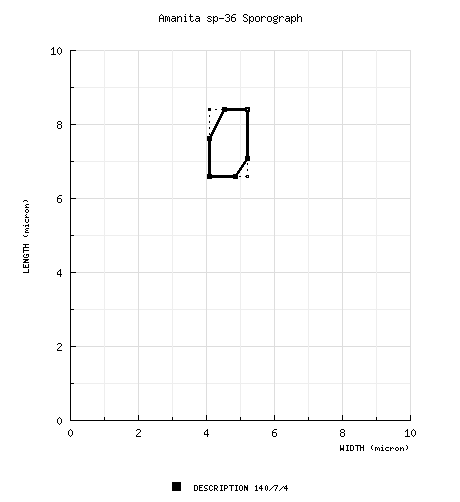
| spores | The spores of this species measure (5.9-) 6.6 - 8.4 (-9.8) × (3.8-) 4.1 - 5.2 (-6.3) μm and are ellipsoid to elongate (infrequently broadly ellipsoid, rarely cylindric) and amyloid. Clamps are lacking at the bases of basidia. |
| discussion |
Amanita sp-36 is known from Oak-Beech-Hickory forest and mixed northern hardwood-conifer forest in the eastern U.S. This is apparently the species to which the name Amanita franchetii was applied by David T. Jenkins (1986). Amanita franchetii is a European taxon.—R. E. Tulloss and N. Goldman |
| brief editors | RET |
| name | Amanita sp-36 | ||||||||
| name status | cryptonomen temporarium | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| intro |
Olive text indicates a specimen that has not been
thoroughly examined (for example, for microscopic details) and marks other places in the text
where data is missing or uncertain. The following material is based upon original research by R. E. Tulloss. RET believes this to be Amanita franchetii sensu Dav. T. Jenkins (1986). | ||||||||
| pileus | 28 - 59± mm wide, orange-brown (ca. 6C8) or ranging from 6D6 near disc to 10YR6/8 to 10YR8/8 (at margin), hemispheric at first with margin straight or slightly decurved, tacky becoming dry, dull; context white, except bright yellow immediately below pileipellis, not staining when cut or bruised, 1 - 4± mm thick over stipe, thinning evenly to margin or thinning to within 2-6 mm of margin and becoming membranous from that point to margin; margin nonstriate, usually nonappendiculate, sometimes bearing fragments of partial veil; universal veil as small warts and confluent warts, orangish yellow to canary yellow to yellow, verruculose, friable, detersile, color change not observed. | ||||||||
| lamellae | very narrowly adnate (10× lens) and lacking decurrent lines on stipe apex, close to crowded, white to pale yellow in mass, color in side view not recorded, unchanging when cut or bruised, 4 - 5.5 mm broad, broadest at mid-length, with yellow edge; lamellulae truncate with attenuate tooth along underside of pileus context, unevenly distributed, of diverse lengths. | ||||||||
| gleba | double click in markup mode to edit. | ||||||||
| stipe | 47 - 84± × 4 - 9.5± mm, white to pale yellow to yellow (variation seen within single collection of three basidiomes), with yellow pigment extensively and evenly distributed or limited to apical or basal portions or only above partial veil, unchanging after cutting or bruising, narrowing upward, flaring at apex, longitudinally striatulate; bulb subglobose, 15± × 14± mm or as clavate stipe base; context white, unchanging when cut or bruised, with central cylinder 1.5 - 3± mm wide, stuffed with white compressed cottony material, larval tunnels sometimes present and then unstaining; partial veil superior to submedian, membranous, thin, persistent, white or yellow above, yellowish below, sometimes becoming gray-brown with age, not striate above, floccose-fibrillose below; universal veil as minute yellow granules or flecks on stipe and loose detersile patches on and in soil surrounding stipe base. | ||||||||
| odor/taste | Odor faintly earthy or absent. Taste not recorded. | ||||||||
| macrochemical tests |
Spot test for tyrosinase: L-tyrosine - positive in lower two-thirds of stipe and in bulb, on cut surfaces of sectioned partial veil and pileipellis, around edges of lamellae (above and below), and at point of merger of stipe and pileus contexts. Test voucher: Tulloss 8-1-86-H. | ||||||||
| basidiospores | RET/CRC: [140/7/4] (5.9-) 6.6 - 8.4 (-9.8) × (3.8-) 4.1 - 5.2 (-6.3) μm, (L = 7.2 - 7.7 μm; L' = 7.5 μm; W = 4.5 - 5.0 μm; W' = 4.8 μm; Q = (1.27-) 1.36 - 1.86 (-2.04); Q = 1.49 - 1.69; Q' = 1.58), hyaline, colorless, smooth, thin-walled, amyloid, ellipsoid to elongate, infrequently broadly ellipsoid, occasionally pip-shaped, sometimes swollen at one end, adaxially flattened; apiculus sublateral, cylindric; contents multiguttulate; white in deposit. | ||||||||
| ecology | In small groups. At 60 - 75 m elev. Connecticut: In dark loam of mixed forest including Pinus, Quercus, Tsuga canadensis, etc. New Jersey: In mature oak-beech-hickory (Quercus-Fagus grandifolia-Carya ovata) forest also including Acer. | ||||||||
| material examined |
U.S.A.: CONNECTICUT—Middlesex Co. - E. Haddam, Devil's Hopyard St. Pk. [41°28’32” N/ 72°20’25” W, 72 m], 4.ix.2011 Josh Hutchins s.n. (RET 489-3, nrITS & nrLSU seq'd.); Salmon River St. For. (South) [41°32’58” N/ 72°27’01” W, 21 m], 3.ix.2011 Paula DeSanto s.n. [Tulloss 9-3-11-K] (RET 490-7, nrITS & nrLSU seq'd.). Unkn. Co. - unkn. loc. 24.viii.2007 COMA2007 foray participant s.n. [Tulloss 8-24-07-T] (RET 438-9, nrITS & nrLSU seq'd.).
NEW JERSEY—Burlington Co. - Brendan T. Byrne | ||||||||
| discussion |
This is apparently the species to which the name A. franchetii was applied by David T. Jenkins (1986). However, A. franchetii is a European taxon, while the present species appears to be endemic to eastern North America. Occasionally this species is mis-identified as A. flavorubens; however the present species generally smaller has different spores size/shape. The following sporograph illustrates the latter observation: Preliminary morphological and genetic data (nrITS and nrLSU) indicate that the species called "sp-F14" in these pages may be very closely related to the present species. RET retains the two temporarily coded entities as separate for the moment on the grounds of difference in spore size and shape. Also, it is possible that A. sp-F14 may be identical to the poorly known A. fraterna. As more information develops, you'll read it here. RET has misidentified older specimens of this species as belonging in A. sp-N47 | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.