


| name | Amanita ingwa |
| name status | nomen acceptum |
| author | Grgur. |
| english name | "Grgurinovic's Dark Amanita" |
| images | |
| intro |
The following is based on the original description by C. A. Grgurinovic (1997). |
| cap |
The cap of Amanita ingwa is up to 60 mm wide, convex when young, becoming almost plane with age, with an entire margin. [Note: Oddly, the color of the cap is not given in the original description. From the name an aboriginal word for "dark," one would assume the cap is dark brown at first.] As it expands, the cap can become virgate. The cap appears velvety over most of the surface at first because it is covered by remnants of the volva. The volva may be somewhat warty in the center. The volva is yellowish at first, becoming browner towards the center; some of it may persist as warts in later development. |
| gills |
The gills are almost free, crowded, and white. The short gills are present in one or two series. |
| stem |
The stem is up to 80 × 18 mm (width measured at apex), white, becoming slightly brownish towards the bulbous base (up to 30 mm wide). |
| spores |
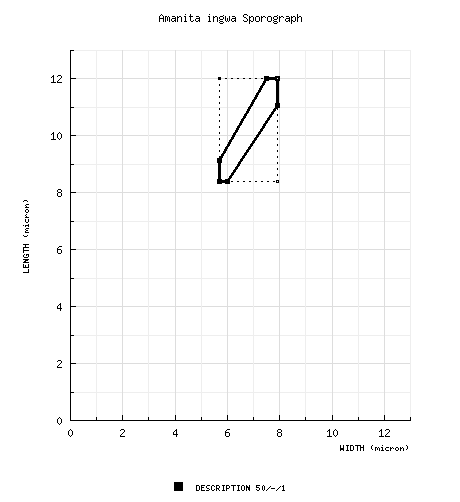
The spores measure 8.4 - 12.0 × 5.7 - 7.9 µm and are ellipsoid to broadly ellipsoid and amyloid. Clamps are absent at bases of basidia. |
| discussion |
Amanita ingwa was originally described from the state of South Australia. No associated plants were mentioned. This species is a bit unusual for Amanita sect. Validae because the limited drawing of the subhymenium in the original description comprises uninflated cells. I cannot recall another species of that section with this character. Perhaps, the drawing doesn't fully represent the mature subhymenium or represents a subhymenium of a specimen too old or poorly preserved to be rehydrated.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita ingwa | ||||||||
| author | Grgur. 1997. Larger Fung. S. Austral.: 404, fig. 269(a-d). | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Grgurinovic's Dark Amanita" | ||||||||
| etymology | ingwa - "dark" (from unidentified language of original peoples of Australia) | ||||||||
| MycoBank nos. | 443364 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | AD | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is derived entirely from the protolog of this species. | ||||||||
| pileus | from protolog: up to 60 mm wide, up to 15 mm high, color not described, dark (per meaning of specific epithet), at length virgate, convex when young, becoming almost plane with age, smooth; context white; margin smooth; universal veil at first forming velvety covering of most of pileus, breaking up into warts, yellowish (4B4 "tinge"), becoming browner (5D5) towards center, in age, colored like pileus surface. | ||||||||
| lamellae | from protolog: almost free, crowded, white, with edge entire; lamellulae in one or two series. | ||||||||
| stipe | from protolog: up to 80 × 18 mm (width measured at apex), white, becoming brownish toward base; bulb 30 mm wide; context not described; partial veil white, membranous, pendulous, faintly striate; universal veil not evident. | ||||||||
| odor/taste | not recorded. | ||||||||
| macrochemical tests |
none recorded. | ||||||||
| pileipellis | not described. | ||||||||
| pileus context | not described. | ||||||||
| lamella trama | not described. | ||||||||
| subhymenium | not described. | ||||||||
| basidia | from protolog: [25/1/1] 33 - 48 × 9.8 - 11.8 μm, with avg. length = 40 μm, with avg. width = 10.7 μm, 4-sterigmate, with sterigmata up to 5.0 μm long; clamps absent. | ||||||||
| universal veil | from protolog: filamentous hyphae [31/1/1] 2.6 - 6.4 μm wide (with avg. width = 4.5 μm), thin-walled, hyaline; inflated cells predominatng, globose or ovoid, 38 - 112 × 15.2 - 71 μm, with avg. length = 72 μm, with avg. width = 39 μm, terminal singly or in short chains (per figure). | ||||||||
| stipe context | not described. | ||||||||
| partial veil | not described. | ||||||||
| lamella edge tissue | from protolog: inflated cells [24/1/1] 5.6 - 27 μm wide (with average width = 17.5 μm), cylindric or clavate or ovoid or spheropedunculate, terminal on short chain of hyphal segments. [Note: This tissue, described by Grgurinovic as "veil fragments," appears to correspond, at least in part, to lamella edge tissue.—ed.] | ||||||||
| basidiospores |
from protolog: [50/-/1] 8.4 - 12.0 × 5.7 - 7.9 μm, (L = 10.2 μm; W = 6.9 μm; Q = 1.50), broadly ellipsoid to ellipsoid, amyloid; apiculus sublateral (per figure); contents not recorded; white in deposit. [Note: We have not estimated a range for Q based on the (Grgurinovic 1997) spore data for this species in order to prevent automatic generation of a sporograph that we believe would be misleading. We evaluated the Amanita spore length and width ranges from (Grgurinovic 1997) in comparison to comparable data published by other authors and often based on revision of the same specimens. This experiment involved a total of 19 descriptions of a total of 13 species from the work of 3 authors. In a range of the form "x - y" of spore length (width) from (Grgurinovic 1997) compared to a range of the form (a-) b - c (-d) of spore length (width) in the other works, the value of "y" was compared to the value of "c" as a ratio. In the case of spore length ranges, on average (per author), the ratio y/c ranged from 1.06 - 1.10 (possibly due to the non-segregation of a "d" value in the ranges of concern). In the case of spore width ranges, on average (per author), the ratio ranged from 1.14 - 1.23 (indicating the probability of compounding causes at play—possibly, the absence of the "d" value in the ranges of concern and failure to restrict spore measurement to spores strictly presenting in lateral view). When sporographs were attempted from the Grgurinovic data, the results were not useful.—ed.] | ||||||||
| ecology | from protolog: Scattered on ground. | ||||||||
| material examined | from protolog: AUSTRALIA: SOUTH AUSTRALIA—Unkn. LGA - Morialta Falls, 28.vi.1984 C. Grgurinovic & R. E. Halling s.n. (holotype, AD 10034). | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.