


| name | Amanita cyanopus |
| name status | nomen acceptum |
| author | C. Simmons, T. Henkel & Bas |
| english name | "Henkel's Blue Foot Amanita" |
| images |
  1. Amanita cyanopus, Pakaraima Mtns., Guyana. |
| intro |
This description is based on the original description (2002). |
| cap |
The cap of Amanita cyanopus is more or less 65 mm wide, plano-concave, rather dull grayish to bluish turquoise, with a smooth margin. The volva is present as flat, felted patches particularly near the margin. |
| gills |
The gills are free, crowded, thickish, occasionally forking, pale cream buff, with a finely uneven, light blue to grayish tan edge. |
| stem |
The stem is 95 × 12 mm, cylindric, annulate, light bluish under a coating of minute, gray, floccose scales. The bulb is slenderly turnip-shaped, 55 × 18 mm, and somewhat rooting, with small, bright blue, conical volval warts and ridges on a paler blue to whitish background. A ring is present at the top of the stem but is lost in the specimen that was photographed. The smell is unpleasant and "distinctly of chlorine." |
| spores |
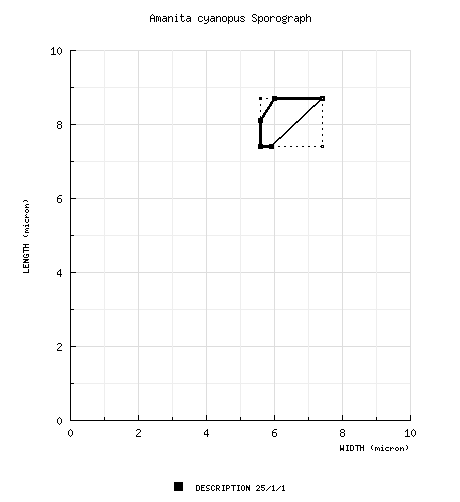
The spores measure 7.4 - 8.7 (-9.0) × (5.0-) 5.6 - 7.4 µm and are broadly ellipsoid to ellipsoid and amyloid. Clamps are absent at the bases of basidia. |
| discussion | The present species grows on an elevated root mat but with hyphae extending down into mineral soil. It was described from a forest dominated by Dicymbe corymbosa and is in the Pakaraima Mountains of Guyana and is known only from the type locality. This is the only species of Amanita known to have a blue pigment. Blue-green staining is known from Amanita pelioma Bas (sect. Lepidella)—an extremely distant relative.—R. E. Tulloss and L. Possiel |
| brief editors | RET |
| name | Amanita cyanopus | ||||||||
| author | C. Simmons, T. Henkel & Bas. 2002. Persoonia 17(4): 569, fig. 2(a-f), pl. 4. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Henkel's Blue Foot Amanita" | ||||||||
| MycoBank nos. | 374037 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | BRG | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is derived from the protolog of the present species. | ||||||||
| basidiospores | from protolog: [25/1/1] 7.4 - 8.7 (-9.0) × (5.0-) 5.6 - 7.4 μm, (Q = (1.15-) 1.25 - 1.45 (-1.60); Q = 1.35), colorless, smooth, thin-walled, amyloid, broadly ellipsoid to ellipsoid; apiculus rather small; contents not described; color in deposit not recorded. | ||||||||
| ecology | "On root mat, but rooting into mineral soil, in forest of predominantly Dicymbe corymbosa (Paluwayek). | ||||||||
| material examined | from protolog: GUYANA: POTARO-SIPARUNI—Pakaraima Mtns., Upper Ireng watershed - W bank of Yuarka R., 1 km upstream from juncture with Suruwabaru Crk., 30.v.1999 T. Henkel et al. TH7083 (holotype, BRG). | ||||||||
| discussion | This species is known only from its type locality. | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.