


| name | Amanita clelandii |
| name status | nomen acceptum |
| author | E.-J. Gilbert |
| english name | "Cleland's Lepidella" |
| intro | This description is largely based on that of Reid (1980), which includes references to the extremely limited original description. |
| cap | The cap of Amanita clelandii is 25 mm wide, slightly convex, and white. |
| gills | The gills just reach the stipe and are a bit crowded and white. |
| stem | The stem is 37 × 8 mm, with oblong cross-section, subhollow, slighty farinose, white, with a submarginate, bulbous base. |
| spores | The spores measure 13 - 16 × 5.5 - 6.8 µm according to the original description or 9.0 - 16.0 × 4.5 - 6.0 µm according to Reid and are broadly ellipsoid and amyloid. Basidia do not bear clamp connections according to Reid. |
| discussion |
This species was originally described from material collected in the state of South Australia, Australia. No habitat information is reported. From the information known about the two species, A. clelandii can be distinguished from A. angustispora Cleland by the presence of basidial clamps in the latter. Macroscopically, the two species are very similar. I am inclined to treat the present species as belonging in sect. Lepidella because, among other things, the present species clearly lacks the robust volva characteristic of sect. Amidella and has a bulbous stipe base.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita clelandii | ||||||||
| author | E.-J. Gilbert. 1941. Iconogr. Mycol. (Milan) 27, suppl. (2): 295. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Cleland's Lepidella" | ||||||||
| synonyms |
≡Amidella clelandii E.-J. Gilbert nom. nud. 1940. Iconogr. Mycol. (Milan) 27, suppl. (1): 77, 130, tab. 25 (figs. 2-3). [Lacking Latin diagnosis, specification of holotype, etc. ICBN §36.1, §37.1, etc.] The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| MycoBank nos. | 284050, 284129 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| lectotypes | AD | ||||||||
| lectotypifications | Reid. 1980. Austral. J. Bot., Suppl. Ser. 8: 18. | ||||||||
| revisions | Reid. 1980. Austral. J. Bot., Suppl. Ser. 8: 18, figs. 4(a-b), 55. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is drawn from the protolog of the present species and from revision of Reid (1980) and Grgurinovic (1997). The history of this species is linked with that of A. angustispora. Among the material sent to him by Cleland and labeled "A. angustispora," Gilbert (1940: 130) found that at least two of these collections bore spores that were of distinctly different shape than those that Gilbert took to be typical of material cited in Cleland's protolog to A. angustispora. Gilbert proposed the present species based on those two collections, designating one collected at Encounter Bay (without a date) as type. As Reid (1980) noted, that collection appears to be lost along with the rest of Gilbert's herbarium. Fortunately, there is material in AD marked, in what Reid took to be ("probably") Gilbert's hand, "Amanita clelandii nov. sp." The date of this collection is 13.ix.1924. We agree with Reid that it is reasonable to assume that this is the second collection seen by Gilbert (although Gilbert apparently mistranscribed the date, which appears as "15.ix.1924" in (Gilbert (loc. cit.). It is this latter collection that Reid proposed as lectotype for the present species. It is now the only known specimen of A. clelandii. | ||||||||
| pileus | from collector's annotation per Reid (1980) re lectotype: 25 mm wide, white, slightly convex, smooth; context not described; margin not described; universal veil not described. | ||||||||
| lamellae | from collector's annotation per Reid (1980) re lectotype: just reaching stipe, close, white; lamellulae not described. | ||||||||
| stipe | from collector's annotation per Reid (1980) re lectotype: 38 mm long, slender, white, slightly mealy; bulb present, slightly(?) marginate; context having "tendency to be hollow"; partial veil not described; universal veil not described. | ||||||||
| odor/taste | not recorded. | ||||||||
| macrochemical tests |
none recorded. | ||||||||
| pileipellis | not described. | ||||||||
| pileus context | not described. | ||||||||
| lamella trama | not described. | ||||||||
| subhymenium | from Reid (1980) re lectotype: clamps "not seen." | ||||||||
| basidia |
from Reid (1980) re lectotype: up to 42 × 11 μm, clavate, "probably immature"; clamps "not seen." from Grgurinovic (1997): [4/-/1] 34 - 44 × 11.2 - 14.0 μm, 4-sterigmate, with sterigmata up to 3.2 μm long; clamps "not seen." | ||||||||
| universal veil | from Reid (1980) re lectotype: On pileus, in inconspicuous felty patches near margin: with elements apparently "irregularly disposed"; hyphae 4 - 10 μm wide, branched, thin-walled, hyaline; inflated cells up to 55 × 30 μm, ovoid or ellipsoid. | ||||||||
| stipe context | not described. | ||||||||
| partial veil | absent? | ||||||||
| lamella edge tissue | not described. | ||||||||
| basidiospores |
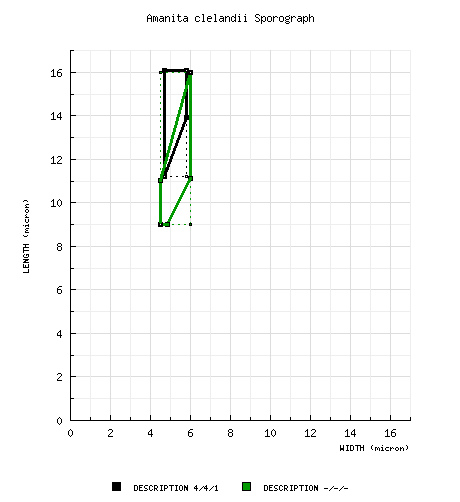
from spore drawings of Gilbert (1940): [4/4/1] 11.2 - 16.1 × 4.7 - 5.8 μm, (L = 14.0 μm; W = 5.2 μm; Q = 2.40 - 3.43; Q = 2.72), hyaline, smooth, not at all amyloid, cylindric to bacilliform; apiculus sublateral (per figure); contents not described; color in deposite not described. [Note: Gilbert, who is responsible for the division of Amanita into two subgenera based on the amyloid reaction of their spores and was well aware that species of sect. Amidella had amyloid spores (Gilbert 1940 & 1941, Bas 1969: 296), wrote in the protolog of the present species both that the spores of the present species were "not at all amyloid" and that its greatest affinity was with A. peckiana, a species that Gilbert knew to have amyloid spores.—ed.] from Reid (1980): [-/-/-] 9.0 - 16.0 × 4.5 - 6.0 μm, (est. Q = 1.85 - 2.45; est. Q = 2.15), "nonamyloid"/amyloid, elongate to cylindric; apiculus sublateral (per figure); contents not described; color in deposit unknown. [Note: Within the space of three short paragraphs, Reid states that the spores of this species are "nonamyloid" and that, in comparing the present species to A. angustispora, the latter is similar to the present species in having "distinctive, amyloid, cylindric spores."—ed.] from Grgurinovic (1997): [50/1/1] 10.0 - 16.0 × 5.2 - 8.0 μm, (L = 13.3 μm; W = 6.5 μm; Q = 2.0), "inamyloid," apparently elongate to cylindric; apiculus sublateral (per figure); contents not described; color in deposit unknown. [Note: Grgurinovic maintains consistently that the spores are inamyloid.—ed.] [Note: We have not estimated a range for Q based on the (Grgurinovic 1997) spore data for this species in order to prevent automatic generation of a sporograph that we believe would be misleading. We evaluated the Amanita spore length and width ranges from (Grgurinovic 1997) in comparison to comparable data published by other authors and often based on revision of the same specimens. This experiment involved a total of 19 descriptions of a total of 13 species from the work of 3 authors. In a range of the form "x - y" of spore length (width) from (Grgurinovic 1997) compared to a range of the form (a-) b - c (-d) of spore length (width) in the other works, the value of "y" was compared to the value of "c" as a ratio. In the case of spore length ranges, on average (per author), the ratio y/c ranged from 1.06 - 1.10 (possibly due to the non-segregation of a "d" value in the ranges of concern). In the case of spore width ranges, on average (per author), the ratio ranged from 1.14 - 1.23 (indicating the probability of compounding causes at play—possibly, the absence of the "d" value in the ranges of concern and failure to restrict spore measurement to spores strictly presenting in lateral view). When sporographs were attempted from the Grgurinovic data, the results were not useful.—ed.] | ||||||||
| ecology | from type study of Reid (1980): . | ||||||||
| material examined | from type studies of Reid (1980) and Grgurinovic (1997): AUSTRALIA: SOUTH AUSTRALIA—Encounter Bay, 13.ix.1924 J. B. Cleland s.n. (lectotype, ADW 3012 => AD 9173). | ||||||||
| discussion |
We note that two authors confused the matter of whether the spores of the present species are amyloid. Reid suggests that the hymenium of the sole collection of A. clelandii is immature. This could certainly have had an impact regarding production of spores that were larger than would be normal in mature material. Also, in species with normally amyloid spores walls, the walls in immature spores are sometimes not as strongly amyloid as in mature ones. From the limited macro- and microscopic evidence available, there does seem to be some significant similarities between the present taxon, A. angustispora, and A. subalbida as Reid points out. It is even possible that, with the availability of future collections of high quality, a case may be made the three names are taxonomic synonyms. Reid emphasizes the view that the spores of the present species are, in some sense, the same as those of A. angustispora. A figure comparing the sporographs of A. angustispora, A. clelandii, and A. subalbida follows: Reid also emphasizes the poor condition of the exsiccata of the types of the three taxa saying that the lectotype of A. clelandii is in worse condition than that of A. angustispora. Bas emphasized that clamps were scarce and hard to discern in A. subalbida. Would clamps be found in new material of the present species? At present, it seems best to keep the three taxa together on a provisional basis. | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita clelandii |
| bottom links |
[ Section Lepidella page. ]
[ Amanita Studies home. ]
[ Keys & Checklists ] |
| name | Amanita clelandii |
| bottom links |
[ Section Lepidella page. ]
[ Amanita Studies home. ]
[ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.